
–°–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А–∞ –Ї–∞–Ї —Б–њ–Њ—Б–Њ–± –≤—Л—П–≤–ї–µ–љ–Є—П –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–Њ–є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є —А–∞—Б—В–µ–љ–Є–є.
–°–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А–∞ –Ї–∞–Ї —Б–њ–Њ—Б–Њ–± –≤—Л—П–≤–ї–µ–љ–Є—П –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ–Њ–є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є —А–∞—Б—В–µ–љ–Є–є.
–Э. –Э. –Я–†–Ю–Ґ–Р–°–Ю–Т–Р
–Ш–љ—Б—В–Є—В—Г—В —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є–Є —А–∞—Б—В–µ–љ–Є–є –Є–Љ. –Ъ. –Р. –Ґ–Є–Љ–Є—А—П–Ј–µ–≤–∞ –Р–Ї–∞–і–µ–Љ–Є–Є –љ–∞—Г–Ї
–°–°–°–†, –Ь–Њ—Б–Ї–≤–∞
–Я—А–Є–Љ–µ–љ—П–µ–Љ—Л–µ —Б–Њ–Ї—А–∞—Й–µ–љ–Є—П:
–Ъ–Я–Ф –ї–∞–Љ–њ—Л - –Њ—В–љ–Њ—И–µ–љ–Є–µ —Н–љ–µ—А–≥–Є–Є –§–Р–† –Ї–Њ –≤—Б–µ–є –њ–Њ—В—А–µ–±–ї—П–µ–Љ–Њ–є —Н–љ–µ—А–≥–Є–Є, –Ъ—Е–Њ–Ј - –Њ—В–љ–Њ—И–µ–љ–Є–µ –њ–Њ–ї–µ–Ј–љ–Њ–є —З–∞—Б—В–Є —Г—А–Њ–ґ–∞—П –Ї –Њ–±—И–µ–є –±–Є–Њ–Љ–∞—Б—Б–µ, –Ь–У–Ы(–Ф–†–Ш) - –Љ–µ—В–∞–ї–ї–Њ–≥–∞–ї–Њ–≥–µ–љ–љ—Л–µ –ї–∞–Љ–њ—Л, –Ф–Э–∞–Ґ - –і—Г–≥–Њ–≤—Л–µ –љ–∞—В—А–Є–µ–≤—Л–µ —В—А—Г–±—З–∞—В—Л–µ –ї–∞–Љ–њ—Л, –§–Р–†- —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Є –∞–Ї—В–Є–≤–љ–∞—П —А–∞–і–Є–∞—Ж–Є—П - –Є–Ј–ї—Г—З–µ–љ–Є–µ –≤ –Њ–±–ї–∞—Б—В–Є 380—720 –љ–Љ, –•–ї - —Е–ї–Њ—А–Њ—Д–Є–ї–ї.
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–ї–Є —А–Њ—Б—В, —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–Є (—Б–∞–ї–∞—В, —А–µ–і–Є—Б, –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї, –њ–µ—А–µ—Ж, –ї–µ–≤–Ј–µ—П —Б–∞—Д–ї–Њ—А–Њ–≤–Є–і–љ–∞—П) –њ—А–Є –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–Є –Є—Е –љ–∞ —Б–≤–µ—В—Г —А–∞–Ј–ї–Є—З–љ–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –Є —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –≤ —Г—Б–ї–Њ–≤–Є—П—Е —Д–Є—В–Њ—В—А–Њ–љ–∞. –Я–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ —Б–≤–µ—В–Њ–≤–Њ–µ –љ–∞—Б—Л—Й–µ–љ–Є–µ –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–Њ—Б—В–∞ –љ–∞—Б—В—Г–њ–∞–µ—В –њ—А–Є –±–Њ–ї–µ–µ –љ–Є–Ј–Ї–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞, —З–µ–Љ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј. –Э–∞–Є–±–Њ–ї–µ–µ –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–Љ–Є –і–ї—П –≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–Є —П–≤–ї—П—О—В—Б—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–† –≤ –њ—А–µ–і–µ–ї–∞—Е 150—220 –Т—В/–Љ2. –°–Є–љ–Є–є —Б–≤–µ—В –≤—Л–Ј—Л–≤–∞–µ—В —В–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ —А–Њ—Б—В–∞ —Б—В–µ–±–ї—П –Є –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –ї–Є—Б—В—М–µ–≤, –њ—А–Є —Н—В–Њ–Љ —Д–Њ—А–Љ–Є—А—Г—О—В—Б—П –ї–Є—Б—В—М—П —Б –±–Њ–ї—М—И–µ–є —Г–і–µ–ї—М–љ–Њ–є –њ–ї–Њ—В–љ–Њ—Б—В—М—О. –Э–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г –љ–∞–±–ї—О–і–∞–ї—Б—П —Б–∞–Љ—Л–є –≤—Л—Б–Њ–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –≤ —А–∞—Б—З–µ—В–µ –љ–∞ –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞. –Ъ—А–∞—Б–љ–∞—П –Њ–±–ї–∞—Б—В—М —Б–њ–µ–Ї—В—А–∞ —Б–њ–Њ—Б–Њ–±—Б—В–≤–Њ–≤–∞–ї–∞ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–Љ—Г —А–Њ—Б—В—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ –Є –≤—Л—В—П–≥–Є–≤–∞–љ–Є—О –Њ—Б–µ–≤—Л—Е –Њ—А–≥–∞–љ–Њ–≤. –Т –Ј–µ–ї–µ–љ–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М —В–Њ–љ–Ї–Є–µ –ї–Є—Б—В—М—П —Б –Љ–µ–љ—М—И–Є–Љ —З–Є—Б–ї–Њ–Љ –Ї–ї–µ—В–Њ–Ї –Є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–Њ–≤ –≤ 1 —Б–Љ2 –ї–Є—Б—В–∞ –Є —А–µ–≥–Є—Б—В—А–Є—А–Њ–≤–∞–ї—Б—П —Б–∞–Љ—Л–є –љ–Є–Ј–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –љ–∞ –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞, –љ–Њ —Б–∞–Љ—Л–є –≤—Л—Б–Њ–Ї–Є–є - –≤ —А–∞—Б—З–µ—В–µ –љ–∞ —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В. –Ш—Б–њ–Њ–ї—М–Ј—Г—П –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л–µ –Є –Љ–µ—В–∞–ї–ї–Њ–≥–∞–ї–Њ–≥–µ–љ–љ—Л–µ –ї–∞–Љ–њ—Л –≤ —Г—Б–ї–Њ–≤–Є—П—Е —Д–Є—В—А–Њ–љ–∞, –њ–Њ–ї—Г—З–µ–љ—Л —Г—А–Њ–ґ–∞–Є –≤ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —А–∞–Ј –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–µ, —З–µ–Љ –≤ —В–µ–њ–ї–Є—Ж–∞—Е –Є –≤ –њ–Њ–ї–µ –≤ 1,5—2 —А–∞–Ј–∞ –±–Њ–ї–µ–µ –Ї–Њ—А–Њ—В–Ї–Є–µ —Б—А–Њ–Ї–Є. –°–і–µ–ї–∞–љ –≤—Л–≤–Њ–і, —З—В–Њ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–µ —Н–љ–µ—А–≥–Є–Є –њ–Њ —Б–њ–µ–Ї—В—А—Г –§–Р–† –≤ —А–∞—Б—В–µ–љ–Є–µ–≤–Њ–і—З–µ—Б–Ї–Є—Е –ї–∞–Љ–њ–∞—Е –ґ–µ–ї–∞—В–µ–ї—М–љ–Њ –Є–Љ–µ—В—М —Б–ї–µ–і—Г—О—Й–µ–µ: 25-30% - –≤ —Б–Є–љ–µ–є (–ї–∞–Љ–њ—Л –Ф–†–Ш), 20% - –≤ –Ј–µ–ї–µ–љ–Њ–є, 50%- –≤ –Ї—А–∞—Б–љ–Њ–є –Њ–±–ї–∞—Б—В–Є (–ї–∞–Љ–њ—Л –Ф–Э–∞–Ґ). –°–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А–∞ - –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М –Є —Б–њ–µ–Ї—В—А —Б–≤–µ—В–∞ - —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј - —Г—А–Њ–ґ–∞–є.
–°–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А–∞ —А–∞—Б—В–µ–љ–Є–є –љ–∞—Е–Њ–і–Є—В –≤—Б–µ –±–Њ–ї–µ–µ —И–Є—А–Њ–Ї–Њ–µ –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ –≤ —Б–µ–ї—М—Б–Ї–Њ—Е–Њ–Ј—П–є—Б—В–≤–µ–љ–љ–Њ–Љ –њ—А–Њ–Є–Ј–≤–Њ–і—Б—В–≤–µ –і–ї—П –і–Њ—Б–≤–µ—З–Є–≤–∞–љ–Є—П —А–∞—Б—Б–∞–і—Л –Њ–≤–Њ—Й–љ—Л—Е –Є –і–µ–Ї–Њ—А–∞—В–Є–≤–љ—Л—Е –Ї—Г–ї—М—В—Г—А –Є –њ—А–Є –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–Є —А–∞—Б—В–µ–љ–Є–є –њ—А–Є –њ–Њ–ї–љ–Њ–Љ –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–Љ –Њ—Б–≤–µ—Й–µ–љ–Є–Є –≤ —В–µ–њ–ї–Є—Ж–∞—Е, –љ–∞ —Б–µ–ї–µ–Ї—Ж–Є–Њ–љ–љ—Л—Е —Б—В–∞–љ—Ж–Є—П—Е –Є —Д–Є—В–Њ—В—А–Њ–љ–∞—Е.
–Т –µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –≤ –њ–µ—А–Є–Њ–і –≤–µ–≥–µ—В–∞—Ж–Є–Є —А–∞—Б—В–µ–љ–Є—П —А–µ–і–Ї–Њ –Є—Б–њ—Л—В—Л–≤–∞—О—В –Њ—Б—В—А—Л–є –љ–µ–і–Њ—Б—В–∞—В–Њ–Ї —Б–Њ–ї–љ–µ—З–љ–Њ–є —А–∞–і–Є–∞—Ж–Є–Є, –Ї–Њ—В–Њ—А—Л–є –њ—А–Є–≤–Њ–і–Є–ї –±—Л –Ї –Є—Е –≥–Є–±–µ–ї–Є. –§–Р–† –Є–Љ–µ–µ—В –Њ—Б–Њ–±—Г—О –Ј–љ–∞—З–Є–Љ–Њ—Б—В—М –≤ —Г—Б–ї–Њ–≤–Є—П—Е –Ј–∞—Й–Є—Й–µ–љ–љ–Њ–≥–Њ –≥—А—Г–љ—В–∞, –≥–і–µ –Є–Ј-–Ј–∞ –љ–Є–Ј–Ї–Њ–є –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є –Є –Ї–Њ—А–Њ—В–Ї–Њ–є –і–ї–Є–љ—Л –і–љ—П –≤ –Њ—Б–µ–љ–љ–µ-–Ј–Є–Љ–љ–Є–µ –Љ–µ—Б—П—Ж—Л –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–µ –њ–Њ–ї–љ–Њ—Ж–µ–љ–љ—Л—Е —А–∞—Б—В–µ–љ–Є–є –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ —В–Њ–ї—М–Ї–Њ —Б –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ–Љ –Є—Б—В–Њ—З–љ–Є–Ї–Њ–≤ –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–≤–µ—В–∞ [1-3].
–Т —Б–≤—П–Ј–Є —Б –љ–µ–Њ–±—Е–Њ–і–Є–Љ–Њ—Б—В—М—О —А–∞—Б—И–Є—А–µ–љ–Є—П –њ–ї–Њ—Й–∞–і–µ–є –њ–Њ–і —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А–Њ–є —А–∞—Б—В–µ–љ–Є–є –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г–≤–µ–ї–Є—З–Є–≤–∞–µ—В—Б—П –њ–Њ—В—А–µ–±–љ–Њ—Б—В—М –≤ –Є—Б—В–Њ—З–љ–Є–Ї–∞—Е –Є–Ј–ї—Г—З–µ–љ–Є—П, –Є–Љ–µ—О—Й–Є—Е –≤—Л—Б–Њ–Ї—Г—О –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї—Г—О —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М. –Ф–ї—П –≤—Л–њ–Њ–ї–љ–µ–љ–Є—П —Н—В–Њ–є –Ј–∞–і–∞—З–Є –љ–µ–Њ–±—Е–Њ–і–Є–Љ –≤—Л–њ—Г—Б–Ї —А–∞—Б—В–µ–љ–Є–µ–≤–Њ–і—З–µ—Б–Ї–Є—Е –ї–∞–Љ–њ, –Є–Љ–µ—О—Й–Є—Е –љ–µ —В–Њ–ї—М–Ї–Њ –≤—Л—Б–Њ–Ї–Є–є –Ъ–Я–Ф, –љ–Њ –Є –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л–є –і–ї—П —А–∞—Б—В–µ–љ–Є–є —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л–є —Б–Њ—Б—В–∞–≤. –Т—Л—Б–Њ–Ї–∞—П –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–∞—П –Є —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–∞—П —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В—М –Є—Б—В–Њ—З–љ–Є–Ї–Њ–≤ –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–≤–µ—В–∞ –≤ –Ї–Њ–љ–µ—З–љ–Њ–Љ —Б—З–µ—В–µ –Њ–њ—А–µ–і–µ–ї–Є—В —Н–Ї–Њ–љ–Њ–Љ–Є—З–µ—Б–Ї–Є–є —Н—Д—Д–µ–Ї—В
—Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л —А–∞—Б—В–µ–љ–Є–є –Є –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В—М –±–Њ–ї–µ–µ —И–Є—А–Њ–Ї–Њ–≥–Њ –µ–µ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є—П –≤ —В–µ–њ–ї–Є—З–љ—Л—Е —Е–Њ–Ј—П–є—Б—В–≤–∞—Е –љ–∞—И–µ–є —Б—В—А–∞–љ—Л.
–Т —Б–≤—П–Ј–Є —Б —В–µ–Љ, —З—В–Њ –і–Њ –љ–∞—Б—В–Њ—П—Й–µ–≥–Њ –≤—А–µ–Љ–µ–љ–Є –≤ –љ–∞—И–µ–є —Б—В—А–∞–љ–µ –љ–µ—В –њ–Њ–ї–љ–Њ—Ж–µ–љ–љ–Њ–≥–Њ —А–∞—Б—В–µ–љ–Є–µ–≤–Њ–і—З–µ—Б–Ї–Њ–≥–Њ –Є—Б—В–Њ—З–љ–Є–Ї–∞ —Б–≤–µ—В–∞, —Б–Њ–Ј–і–∞–љ–Є–µ —В–∞–Ї–Њ–≥–Њ –Є—Б—В–Њ—З–љ–Є–Ї–∞ —П–≤–ї—П–µ—В—Б—П –≤–∞–ґ–љ–Њ–є –Ј–∞–і–∞—З–µ–є –і–ї—П –і–∞–ї—М–љ–µ–є—И–µ–≥–Њ —А–∞–Ј–≤–Є—В–Є—П —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л —А–∞—Б—В–µ–љ–Є–Є. –Т —Н—В–Њ–Љ –њ–ї–∞–љ–µ –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В –Є–љ—В–µ—А–µ—Б —А–∞–±–Њ—В—Л –њ–Њ –Є–Ј—Г—З–µ–љ–Є—О —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ —Б–≤–µ—В–∞, –љ–∞–Є–±–Њ–ї–µ–µ —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ–≥–Њ –і–ї—П –≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П —А–∞—Б—В–µ–љ–Є–є.
–Я–µ—А–µ–і –љ–∞–Љ–Є —Б—В–Њ—П–ї–Є –Ј–∞–і–∞—З–Є –≤—Л—П—Б–љ–Є—В—М:
1) –Ї–∞–Ї–Є–µ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –±–µ–ї–Њ–≥–Њ —Б–≤–µ—В–∞ –≤ —Г—Б–ї–Њ–≤–Є—П—Е —Д–Є—В–Њ—В—А–Њ–љ–∞ —П–≤–ї—П—О—В—Б—П –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л–Љ–Є –Є –Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ –і–Њ–њ—Г—Б—В–Є–Љ—Л–Љ–Є –і–ї—П –≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П —А–∞—Б—В–µ–љ–Є–є;
2) –Ї–∞–Ї–Њ–≤–∞ —Б–њ–µ—Ж–Є—Д–Є—З–љ–Њ—Б—В—М –і–µ–є—Б—В–≤–Є—П –љ–∞ —А–Њ—Б—В –Є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј —В—А–µ—Е –Њ—Б–љ–Њ–≤–љ—Л—Е –Њ–±–ї–∞—Б—В–µ–є –§–Р–† - —Б–Є–љ–µ–є, –Ї—А–∞—Б–љ–Њ–є –Є –Ј–µ–ї–µ–љ–Њ–є;
3) –Ї–∞–Ї –Љ–µ–љ—П–µ—В—Б—П –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–є –≤ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ —Б–≤–µ—В–∞;
4) –Ї–∞–Ї–Њ–µ —А–∞—Б–њ—А–µ–і–µ–ї–µ–љ–Є–µ —Н–љ–µ—А–≥–Є–Є –њ–Њ —Б–њ–µ–Ї—В—А—Г –§–Р–† –ґ–µ–ї–∞—В–µ–ї—М–љ–Њ –Є–Љ–µ—В—М –≤ –Є—Б—В–Њ—З–љ–Є–Ї–µ —Б–≤–µ—В–∞ –і–ї—П —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л —А–∞—Б—В–µ–љ–Є–є.
–£—З–Є—В—Л–≤–∞—П —В–Њ –Њ–±—Б—В–Њ—П—В–µ–ї—М—Б—В–≤–Њ, —З—В–Њ –≤ –µ—Б—В–µ—Б—В–≤–µ–љ–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –њ—А–Њ–Є–Ј—А–∞—Б—В–∞–љ–Є—П —А–∞—Б—В–µ–љ–Є—П –љ–Є–Ї–Њ–≥–і–∞ –љ–µ –њ–Њ–ї—Г—З–∞—О—В —З–Є—Б—В–Њ –Ї—А–∞—Б–љ—Л–є, —З–Є—Б—В–Њ —Б–Є–љ–Є–є –Є–ї–Є —З–Є—Б—В–Њ –Ј–µ–ї–µ–љ—Л–є —Б–≤–µ—В, –∞ –њ–Њ–ї—Г—З–∞—О—В –Є—Е —Б—Г–Љ–Љ—Г —Б —А–∞–Ј–љ—Л–Љ–Є –Љ–∞–Ї—Б–Є–Љ—Г–Љ–∞–Љ–Є –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В –њ–ї–Њ—В–љ–Њ—Б—В–Є –њ–Њ—Б–µ–≤–∞ –Є –≥–µ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–є —И–Є—А–Њ—В—Л –Љ–µ—Б—В–∞, –Љ—Л –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є –Є—Б—В–Њ—З–љ–Є–Ї–Є, –Є–Љ–µ—О—Й–Є–µ –Љ–∞–Ї—Б–Є–Љ—Г–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ —В–Њ–є –Є–ї–Є –і—А—Г–≥–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞, - —Ж–≤–µ—В–љ—Л–µ –ї—О–Љ–Є–љ–µ—Б—Ж–µ–љ—В–љ—Л–µ –ї–∞–Љ–њ—Л.
–Я—А–Є –Њ–њ—А–µ–і–µ–ї–µ–љ–Є–Є –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–є —Н—Д—Д–µ–Ї—В–Є–≤–љ–Њ—Б—В–Є –Є—Б—В–Њ—З–љ–Є–Ї–∞ –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–≥–Њ —Б–≤–µ—В–∞ –Є–ї–Є –Њ—В–і–µ–ї—М–љ—Л—Е –Њ–±–ї–∞—Б—В–µ–є —Б–њ–µ–Ї—В—А–∞ –≤ –Ї–∞—З–µ—Б—В–≤–µ –Њ—Б–љ–Њ–≤–љ—Л—Е –Ї—А–Є—В–µ—А–Є–µ–≤ –Њ—Ж–µ–љ–Ї–Є –Љ—Л –±—А–∞–ї–Є –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –Є —Б–Ї–Њ—А–Њ—Б—В–Є —А–Њ—Б—В–∞ — –Њ—Б–љ–Њ–≤–љ—Л–µ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –њ—А–Њ—Ж–µ—Б—Б—Л, –Њ–њ—А–µ–і–µ–ї—П—О—Й–Є–µ –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–є.
–Ь–µ—В–Њ–і–Є–Ї–∞
–Ю–њ—Л—В—Л –њ—А–Њ–≤–Њ–і–Є–ї–Є –≤ –Ї–Њ–љ–і–Є—Ж–Є–Њ–љ–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ї–∞–Љ–µ—А–∞—Е —Д–Є—В–Њ—В—А–Њ–љ–∞ –Ш–љ—Б—В–Є—В—Г—В–∞ —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є–Є —А–∞—Б—В–µ–љ–Є–є –Р–Э –°–°–°–†. –Т –Ї–∞—З–µ—Б—В–≤–µ –Њ–њ—Л—В–љ—Л—Е —А–∞—Б—В–µ–љ–Є–є –±—Л–ї–Є –≤–Ј—П—В—Л: —Б–∞–ї–∞—В (–ђ–∞—Б–®—Б–∞ –Ј–∞–Э—Г–∞ –ђ), —А–µ–і–Є—Б (–Ъ–∞—А–ђ–∞–њ–Є–Ј –Ј–∞–є—Г–Є–Ј –ђ., —Г–∞–≥. —В—В–Њ–≥), –њ–µ—А–µ—Ж —Б–ї–∞–і–Ї–Є–є (–°–∞—А–Ј—О–Є—В –∞–њ–њ–Є–Є—В –ђ), –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї. (–Э–µ–Э–∞–њ–®–Є–Ј –∞–њ–њ–Є–Є–Ј –ђ.) –Є –ї–µ–≤–Ј–µ—П —Б–∞—Д–ї–Њ—А–Њ–≤–Є–і–љ–∞—П (–Ъ–Ш–∞—А–Њ–њ–Э—Б–Є—В —Б–∞–≥–®–∞—В–Њ–Ђ–µ–Ј '–Ь–Ђ I). –†–∞—Б—В–µ–љ–Є—П –≤—Л—А–∞—Й–Є–≤–∞–ї–Є –≤ –њ–Њ—З–≤–µ–љ–љ–Њ–є –Ї—Г–ї—М—В—Г—А–µ –њ—А–Є 23° –і–љ–µ–Љ –Є 20° –љ–Њ—З—М—О, –≤–ї–∞–ґ–љ–Њ—Б—В–Є –≤–Њ–Ј–і—Г—Е–∞ 75—80% –њ—А–Є —А–∞–Ј–ї–Є—З–љ–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–† –Є —Б–≤–µ—В–Њ–≤–Њ–Љ –і–љ–µ 16 —З. –Ш–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М —Б–≤–µ—В–∞ (–Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В—М) –Є–Ј–Љ–µ—А—П–ї–∞—Б—М –≤ –Т—В/–Љ2 –§–Р–†.
–Ш—Б—В–Њ—З–љ–Є–Ї–Њ–Љ —Б–≤–µ—В–∞ —Б–ї—Г–ґ–Є–ї–Є –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л–µ –ї–∞–Љ–њ—Л –Љ–Њ—Й–љ–Њ—Б—В—М—О 20 –Ї–Т—В –Љ–∞—А–Ї–Є –Ф–Ъ–°–Ґ-20 000 —Б –≤–Њ–і–љ—Л–Љ —Н–Ї—А–∞–љ–Њ–Љ. –Ъ—Б–µ–љ–Њ–љ–Њ–≤—Л–µ –ї–∞–Љ–њ—Л –њ–Њ —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–є —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–µ –≤ –Њ–±–ї–∞—Б—В–Є –§–Р–† –±–ї–Є–Ј–Ї–Є –Ї —Б–Њ–ї–љ–µ—З–љ–Њ–Љ—Г —Б–њ–µ–Ї—В—А—Г. –С–Њ–ї—М—И–∞—П –Љ–Њ—Й–љ–Њ—Б—В—М –ї–∞–Љ–њ –њ–Њ–Ј–≤–Њ–ї–Є–ї–∞ –њ–Њ–ї—Г—З–∞—В—М –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В—М, —А–∞–≤–љ—Г—О –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–є —Б–Њ–ї–љ–µ—З–љ–Њ–є - 500 –Т—В/–Љ2 –§–Р–† (~100 —В—Л—Б. –ї–Ї). –Ш—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є —В–∞–Ї–ґ–µ —Ж–≤–µ—В–љ—Л–µ –ї—О–Љ–Є–љ–µ—Б—Ж–µ–љ—В–љ—Л–µ –ї–∞–Љ–њ—Л —Б –Љ–∞–Ї—Б–Є–Љ—Г–Љ–Њ–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ –Ї—А–∞—Б–љ–Њ–є, —Б–Є–љ–µ–є –Є–ї–Є –Ј–µ–ї–µ–љ–Њ–є –Њ–±–ї–∞—Б—В—П—Е —Б–њ–µ–Ї—В—А–∞: –ї–∞–Љ–њ—Л –Ї—А–∞—Б–љ–Њ–≥–Њ —Б–≤–µ—В–∞ –Ы–Ъ-65, –Є–Ј–ї—Г—З–µ–љ–Є–µ –≤ –Њ–±–ї–∞—Б—В–Є 600-710 –љ–Љ —Б –Љ–∞–Ї—Б–Є–Љ—Г–Љ–Њ–Љ –њ—А–Є 640-660 –љ–Љ –њ–ї—О—Б 14% –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ —Б–Є–љ–µ–є –Њ–±–ї–∞—Б—В–Є; –ї–∞–Љ–њ—Л —Б–Є–љ–µ–≥–Њ —Б–≤–µ—В–∞ –Ы–°-150, –Є–Ј–ї—Г—З–µ–љ–Є–µ –≤ –Њ–±–ї–∞—Б—В–Є 380-600 –љ–Љ —Б –Љ–∞–Ї—Б–Є–Љ—Г–Љ–Њ–Љ –њ—А–Є 440-460 –љ–Љ; –ї–∞–Љ–њ—Л –Ј–µ–ї–µ–љ–Њ–≥–Њ —Б–≤–µ—В–∞ –Ы–Ч-150, –Є–Ј–ї—Г—З–µ–љ–Є–µ –≤ –Њ–±–ї–∞—Б—В–Є 490-605 –љ–Љ —Б –Љ–∞–Ї—Б–Є–Љ—Г–Љ–Њ–Љ –њ—А–Є 520-550 –љ–Љ; —Д–Є—В–Њ–ї–∞–Љ–њ—Л –Ы–§–†-150, –Є–Ј–ї—Г—З–µ–љ–Є–µ –≤ –Њ–±–ї–∞—Б—В–Є 590-720 –љ–Љ —Б –Љ–∞–Ї—Б–Є–Љ—Г–Љ–Њ–Љ –њ—А–Є 600-620 –Љ–Љ (–ї–∞–Љ–њ—Л –Ї—А–∞—Б–љ–Њ–≥–Њ —Б–≤–µ—В–∞ —Б –і–Њ–±–∞–≤–Ї–Њ–є 25% —Б–Є–љ–µ–≥–Њ).
–°–≤–µ—В –±—Л–ї –≤—Л—А–∞–≤–љ–µ–љ –њ–Њ —З–Є—Б–ї—Г –њ–∞–і–∞—О—Й–Є—Е –Ї–≤–∞–љ—В–Њ–≤ –Є —Б–Њ—Б—В–∞–≤–ї—П–ї 19,69*1015 –Ї–≤–∞–љ—В* —Б–Љ-2* —Б-1 (65-83 –Т—В/–Љ2 –§–Р–†). –≠–Ї—Б—В—А–∞–Ї—Ж–Є—О —Е–ї–Њ—А–Њ—Д–Є–ї–ї–Њ–≤ –њ—А–Њ–≤–Њ–і–Є–ї–Є —Б –њ—А–Є–Љ–µ–љ–µ–љ–Є–µ–Љ –Њ—Е–ї–∞–ґ–і–µ–љ–љ–Њ–≥–Њ 85%-–љ–Њ–≥–Њ –∞—Ж–µ—В–Њ–љ–∞. –Ш–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –Њ–њ—А–µ–і–µ–ї—П–ї–Є –њ–Њ –Є–Ј–Љ–µ–љ–µ–љ–Є—О –Ї–Њ–љ—Ж–µ–љ—В—А–∞—Ж–Є–Є –°–Ю2 –≤ –Ј–∞–Љ–Ї–љ—Г—В–Њ–є —Б–Є—Б—В–µ–Љ–µ, —Б–Њ–µ–і–Є–љ–µ–љ–љ–Њ–є —Б –Є–љ—Д—А–∞–Ї—А–∞—Б–љ—Л–Љ –≥–∞–Ј–Њ–∞–љ–∞–ї–Є–Ј–∞—В–Њ—А–Њ–Љ «1–њ–У–≥–∞1—Г1-3» («1–Є–њ–Ї–∞1–Њ–≥», –У–Ф–†). –Ю–њ—А–µ–і–µ–ї–µ–љ–Є—П –њ—А–Њ–≤–Њ–і–Є–ї–Є –љ–∞ –ї–Є—Б—В—М—П—Е, –љ–µ –Њ—В–і–µ–ї–µ–љ–љ—Л—Е –Њ—В —А–∞—Б—В–µ–љ–Є–є —Б –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–љ–Є–µ–Љ –Ї–∞–Љ–µ—А—Л-—Й–Є–њ—Ж–Њ–≤. –°–≤–µ—В–Њ–≤—Л–µ –Ї—А–Є–≤—Л–µ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –њ–Њ–ї—Г—З–∞–ї–Є –њ—А–Є 0,04% –°–Ю2 , –њ–Њ—В–µ–љ—Ж–Є–∞–ї—М–љ—Л–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј - –њ—А–Є –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є 500 –Т—В/–Љ- –§–Р–† –Є 0,08% –°–Ю2 . –Я–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤ –Њ–њ—А–µ–і–µ–ї—П–ї–Є –≤–µ—Б–Њ–≤—Л–Љ –Љ–µ—В–Њ–і–Њ–Љ. –Я–Њ–≥–ї–Њ—Й–µ–љ–Є–µ –Є –Њ—В—А–∞–ґ–µ–љ–Є–µ –§–Р–† –ї–Є—Б—В–Њ–Љ –Њ–њ—А–µ–і–µ–ї—П–ї–Є –љ–∞ —Б–њ–µ–Ї—В—А–Њ—Д–Њ—В–Њ–Љ–µ—В—А–µ –°–§-14 (–°–°–°–†). –Т —В–∞–±–ї–Є—Ж–∞—Е –Є –љ–∞ —А–Є—Б—Г–љ–Ї–∞—Е –њ—А–Є–≤–µ–і–µ–љ—Л —Б—А–µ–і–љ–Є–µ –∞—А–Є—Д–Љ–µ—В–Є—З–µ—Б–Ї–Є–µ –Є–Ј 4-5 –±–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –њ–Њ–≤—В–Њ—А–љ–Њ—Б—В–µ–є –Є –Є—Е —Б—В–∞–љ–і–∞—А—В–љ—Л–µ –Њ—И–Є–±–Ї–Є.
 —Г —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –љ–∞ —Б–≤–µ—В—Г –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л—Е –ї–∞–Љ–њ (1) –Є –њ—А–Є –µ—Б—В–µ—Б—В–≤–µ–љ–љ–Њ–Љ –Њ—Б–≤–µ—Й–µ–љ–Є–Є (2). –†–Є—Б. 2. –Ч–∞–≤–Є—Б–Є–Љ–Њ—Б—В—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ (1) –Є —А–Њ—Б—В–∞ –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ (2) –Њ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞ —Г —А–∞—Б—В–µ–љ–Є–є —А–µ–і–Є—Б–∞. –Ч–∞—И—В—А–Є—Е–Њ–≤–∞–љ–љ–∞—П –Њ–±–ї–∞—Б—В—М - –і–Є–∞–њ–∞–Ј–Њ–љ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є, –љ–∞–Є–±–Њ–ї–µ–µ –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л—Е –і–ї—П –≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є")
–†–Є—Б. 1. –°—Г—В–Њ—З–љ–Њ–µ —Г—Б–≤–Њ–µ–љ–Є–µ –°–Ю2 (–≤ —А–∞—Б—З–µ—В–µ –љ–∞ –Ї–≤.–Љ –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤) —Г —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –љ–∞ —Б–≤–µ—В—Г –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л—Е –ї–∞–Љ–њ (1) –Є –њ—А–Є –µ—Б—В–µ—Б—В–≤–µ–љ–љ–Њ–Љ –Њ—Б–≤–µ—Й–µ–љ–Є–Є (2).
–†–Є—Б. 2. –Ч–∞–≤–Є—Б–Є–Љ–Њ—Б—В—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ (1) –Є —А–Њ—Б—В–∞ –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ (2) –Њ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞ —Г —А–∞—Б—В–µ–љ–Є–є —А–µ–і–Є—Б–∞.
–Ч–∞—И—В—А–Є—Е–Њ–≤–∞–љ–љ–∞—П –Њ–±–ї–∞—Б—В—М - –і–Є–∞–њ–∞–Ј–Њ–љ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є, –љ–∞–Є–±–Њ–ї–µ–µ –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л—Е –і–ї—П
–≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є.
–†–µ–Ј—Г–ї—М—В–∞—В—Л –Є –Њ–±—Б—Г–ґ–і–µ–љ–Є—П
–§–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є —А–Њ—Б—В - –і–≤–∞ –Њ—Б–љ–Њ–≤–љ—Л—Е –њ—А–Њ—Ж–µ—Б—Б–∞ –ґ–Є–Ј–љ–µ–і–µ—П—В–µ–ї—М–љ–Њ—Б—В–Є —А–∞—Б—В–µ–љ–Є–є, –Њ—В —Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ–љ–Њ—Б—В–Є –Ї–Њ—В–Њ—А—Л—Е –Ј–∞–≤–Є—Б–Є—В –Ї–Њ–љ–µ—З–љ—Л–є —Г—А–Њ–ґ–∞–є —А–∞—Б—В–µ–љ–Є–є [4]. –Ю–±–∞ —Н—В–Є –њ—А–Њ—Ж–µ—Б—Б–∞ –≤ —Б–≤–Њ—О –Њ—З–µ—А–µ–і—М –љ–∞—Е–Њ–і—П—В—Б—П –≤ —В–µ—Б–љ–Њ–є –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В–Є –Њ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –Є —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ —Б–≤–µ—В–∞ [5, 6]. –Ш–Ј–≤–µ—Б—В–љ–Њ, —З—В–Њ –≤ –±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –Њ—Б–≤–µ—Й–µ–љ–Є—П –Є –Љ–Є–љ–µ—А–∞–ї—М–љ–Њ–≥–Њ –њ–Є—В–∞–љ–Є—П —Г —А–∞—Б—В–µ–љ–Є–є —Д–Њ—А–Љ–Є—А—Г–µ—В—Б—П –ї–Є—Б—В–Њ–≤–∞—П –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В—М, —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–∞—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М –Ї–Њ—В–Њ—А–Њ–є —Б–њ–Њ—Б–Њ–±–љ–∞ –њ–Њ–ї–љ–Њ—Б—В—М—О –Њ–±–µ—Б–њ–µ—З–Є—В—М —А–Њ—Б—В–Њ–≤—Г—О —Д—Г–љ–Ї—Ж–Є—О –Є —А–µ–њ—А–Њ–і—Г–Ї—В–Є–≤–љ—Л–µ –њ—А–Њ—Ж–µ—Б—Б—Л, –Ј–∞–Ї–Њ–і–Є—А–Њ–≤–∞–љ–љ—Л–µ –≤ –≥–µ–љ–Њ–Љ–µ. –Т –љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л—Е —Б–≤–µ—В–Њ–≤—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е —Г —А–∞—Б—В–µ–љ–Є–є –≤ –њ—А–Њ—Ж–µ—Б—Б–µ —Н–≤–Њ–ї—О—Ж–Є–Є –≤—Л—А–∞–±–Њ—В–∞–ї–Є—Б—М –Ї–Њ–Љ–њ–µ–љ—Б–∞—В–Њ—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л, –≤–Њ—Б–њ–Њ–ї–љ—П—О—Й–Є–µ –љ–µ–і–Њ—Б—В–∞—В–Њ–Ї –Њ–і–љ–Њ–≥–Њ –њ–∞—А–∞–Љ–µ—В—А–∞ –і—А—Г–≥–Є–Љ [7, 8]. –Ґ–∞–Ї, –њ—А–Є –љ–Є–Ј–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ –љ–µ–≤—Л—Б–Њ–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј —З–∞—Б—В–Є—З–љ–Њ –Ї–Њ–Љ–њ–µ–љ—Б–Є—А—Г–µ—В—Б—П —Г—Б–Є–ї–µ–љ–љ—Л–Љ —А–Њ—Б—В–Њ–Љ –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤, –≤ —В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї –њ—А–Є –≤—Л—Б–Њ–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ –Љ–µ–љ—М—И–∞—П –ї–Є—Б—В–Њ–≤–∞—П –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В—М –Љ–Њ–ґ–µ—В –Ї–Њ–Љ–њ–µ–љ—Б–Є—А–Њ–≤–∞—В—М—Б—П –њ–Њ–≤—Л—И–µ–љ–љ–Њ–є —Б–Ї–Њ—А–Њ—Б—В—М—О –∞—Б—Б–Є–Љ–Є–ї—П—Ж–Є–Є –°–Ю2 . –Т –Є—В–Њ–≥–µ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —Г–ї—Г—З—И–∞–µ—В—Б—П –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ—Л–є –±–∞–ї–∞–љ—Б –њ–Њ–≥–ї–Њ—Й–µ–љ–љ–Њ–є —Н–љ–µ—А–≥–Є–Є –Ї–∞–Ї —Г —В–µ–љ–µ–≤—Л—Е, —В–∞–Ї –Є —Г —Б–≤–µ—В–Њ–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є.
–Ч–∞—Б–ї—Г–ґ–Є–≤–∞–µ—В –≤–љ–Є–Љ–∞–љ–Є—П —В–Њ—В —Д–∞–Ї—В, —З—В–Њ –≤ —Г—Б–ї–Њ–≤–Є—П—Е —Д–Є—В–Њ—В—А–Њ–љ–∞, –Ї–Њ–≥–і–∞ –њ–Њ–і–Њ–±—А–∞–љ –Ї–Њ–Љ–њ–ї–µ–Ї—Б –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л—Е —Д–∞–Ї—В–Њ—А–Њ–≤ (—Б–≤–µ—В–Њ–≤–Њ–є –Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–љ—Л–µ —А–µ–ґ–Є–Љ—Л, –Љ–Є–љ–µ—А–∞–ї—М–љ–Њ–µ –њ–Є—В–∞–љ–Є–µ –Є –њ—А.), —А–∞—Б—В–µ–љ–Є—П –Є–Љ–µ—О—В –≤—Л—Б–Њ–Ї–Є–є —Г—А–Њ–≤–µ–љ—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –≤ —В–µ—З–µ–љ–Є–µ –≤—Б–µ–≥–Њ —Б–≤–µ—В–Њ–≤–Њ–≥–Њ –і–љ—П –±–µ–Ј —Б–њ–∞–і–Њ–≤ –Є –њ–Њ–ї—Г–і–µ–љ–љ—Л—Е –і–µ–њ—А–µ—Б—Б–Є–є (—А–Є—Б. 1). –Э–∞ —А–Є—Б. 1 –њ–Њ–Ї–∞–Ј–∞–љ –і–љ–µ–≤–љ–Њ–є —Е–Њ–і —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –≤ —Г—Б–ї–Њ–≤–Є—П—Е –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–≥–Њ –Є –µ—Б—В–µ—Б—В–≤–µ–љ–љ–Њ–≥–Њ –Њ—Б–≤–µ—Й–µ–љ–Є—П. –Э–∞ –≥—А–∞—Д–Є–Ї–µ –≤–Є–і–љ–Њ, —З—В–Њ —Б—Г—В–Њ—З–љ–Њ–µ —Г—Б–≤–Њ–µ–љ–Є–µ —Г–≥–ї–µ–Ї–Є—Б–ї–Њ—В—Л –≤ —А–∞—Б—З–µ—В–µ –љ–∞ 1 –Љ2 –ї–Є—Б—В–∞ —Г —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і —Б–Њ–ї–љ–µ—З–љ—Л–Љ —Б–≤–µ—В–Њ–Љ –њ–Њ—З—В–Є –≤–і–≤–Њ–µ –Љ–µ–љ—М—И–µ, —З–µ–Љ –њ–Њ–і —Б–≤–µ—В–Њ–Љ –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л—Е –ї–∞–Љ–њ.–≠–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П, –њ—А–Њ–≤–µ–і–µ–љ–љ—Л–µ –≤ —Г—Б–ї–Њ–≤–Є—П—Е —Д–Є—В–Њ—В—А–Њ–љ–∞ [5, 6], –њ–Њ–Ј–≤–Њ–ї–Є–ї–Є —Г—Б—В–∞–љ–Њ–≤–Є—В—М, —З—В–Њ –њ—А–Є –і–ї–Є—В–µ–ї—М–љ–Њ–Љ –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–Є —А–∞—Б—В–µ–љ–Є–є –љ–∞ —А–∞–Ј–ї–Є—З–љ—Л—Е –Є–љ—В–µ–њ—Б–љ–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ –≤–њ–ї–Њ—В—М –і–Њ –љ–∞—Б—Л—Й–∞—О—Й–Є—Е, —А–∞–≤–љ—Л—Е –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л–Љ —Б–Њ–ї–љ–µ—З–љ—Л–Љ -500 –Т—В/–Љ2 –§–Р–†, –њ—А–Є 16-—З —Д–Њ—В–Њ–њ–µ—А–Є–Њ–і–µ –њ–Њ –Љ–µ—А–µ –≤–Њ–Ј—А–∞—Б—В–∞–љ–Є—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞ —Г–≤–µ–ї–Є—З–Є–≤–∞—О—В—Б—П —Б–Ї–Њ—А–Њ—Б—В—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –Є —А–∞–Ј–Љ–µ—А –ї–Є—Б—В–Њ–≤–Њ–є –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є (—А–Є—Б. 2). –Ю–і–љ–∞–Ї–Њ –њ—А–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е, –њ—А–µ–≤—Л—И–∞—О—Й–Є—Е 250 –Т—В/–Љ2 –§–Р–†, —В–Њ—А–Љ–Њ–Ј–Є—В—Б—П —А–Њ—Б—В –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ –Є –њ–Њ–і–∞–≤–ї—П–µ—В—Б—П —А–Њ—Б—В —Б—В–µ–±–ї—П (—А–Є—Б. 2). –Я—А–Є —Н—В–Њ–Љ —Б–≤–µ—В –≤—Л—Б–Њ–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є (—Б–≤—Л—И–µ 400 –Т—В/–Љ2 –§–Р–†) –љ–∞—Б—В–Њ–ї—М–Ї–Њ –њ–Њ–і–∞–≤–ї—П–µ—В —А–Њ—Б—В —А–∞—Б—В–µ–љ–Є–Є, —З—В–Њ –≤ —Н—В–Є—Е —Г—Б–ї–Њ–≤–Є—П—Е –≤—Л—А–∞—Б—В–∞—О—В —А–∞—Б—В–µ–љ–Є—П –Ї–∞—А–ї–Є–Ї–Њ–≤–Њ–є —Д–Њ—А–Љ—Л. –Т –Њ—В–ї–Є—З–Є–µ –Њ—В –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є—Е –Ї–∞—А–ї–Є–Ї–Њ–≤ –Љ—Л –Є—Е –љ–∞–Ј–≤–∞–ї–Є «—Б–≤–µ—В–Њ–≤—Л–µ –Ї–∞—А–ї–Є–Ї–Є». –≠—В–Є —Б–≤–µ—В–Њ–≤—Л–µ, –Є–ї–Є —Д–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ, –Ї–∞—А–ї–Є–Ї–Є –Є–Љ–µ–ї–Є –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –Љ–µ–љ—М—И—Г—О –±–Є–Њ–Љ–∞—Б—Б—Г –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б —А–∞—Б—В–µ–љ–Є—П–Љ–Є, –≤—Л—А–∞—Й–µ–љ–љ—Л–Љ–Є –њ—А–Є –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞.
: 30 (1), 80 (2), 200 (3) –Є 420 (4)")
–†–Є—Б. 3. –†–∞—Б—В–µ–љ–Є—П —Б–∞–ї–∞—В–∞, –≤—Л—А–∞—Й–µ–љ–љ—Л–µ –њ—А–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ (–Т—В/–Љ2 –§–Р–†): 30 (1), 80 (2), 200 (3) –Є 420 (4)
–Ґ–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ —А–Њ—Б—В–Њ–≤—Л—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —Б–≤–µ—В–Њ–Љ –≤—Л—Б–Њ–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –Ї–∞–Ї —Г —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е (–њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї, —А–µ–і–Є—Б –Є –і—А.) —В–∞–Ї –Є —Г —В–µ–љ–µ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є (—Б–∞–ї–∞—В –Ь–Њ—Б–Ї–Њ–≤—Б–Ї–Є–є –њ–∞—А–љ–Є–Ї–Њ–≤—Л–є). –°–≤–µ—В–Њ–≤–Њ–µ –љ–∞—Б—Л—Й–µ–љ–Є–µ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ —Г —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є –њ—А–Є –і–ї–Є—В–µ–ї—М–љ–Њ–Љ –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–Є –љ–∞ –≤—Л—Б–Њ–Ї–Њ–Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–Љ —Б–≤–µ—В—Г –љ–∞—Б—В—Г–њ–∞–ї–Њ –њ—А–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–† —Б–≤—Л—И–µ 420 –Т—В/–Љ2. –£ —В–µ–љ–µ–≤—Л–љ–Њ—Б–ї–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–µ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М –њ—А–Є –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В—П—Е –Њ–Ї–Њ–ї–Њ 400 –Т—В/–Љ2, –Њ–љ–Њ —П–≤–ї—П–ї–Њ—Б—М —Б–ї–µ–і—Б—В–≤–Є–µ–Љ —А–∞–Ј—А—Г—И–µ–љ–Є—П –•–ї, –∞ –њ—А–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤—Л—И–µ 420 –Т—В/–Љ2 –§–Р–† - —В–∞–Ї–ґ–µ –Є –і–µ—Б—В—А—Г–Ї—Ж–Є–Є –њ–ї–∞—Б—В–Є–і [6]. 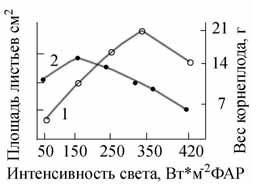
–†–Є—Б. 4. –Ч–∞–≤–Є—Б–Є–Љ–Њ—Б—В—М –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –≤–µ—Б–∞ –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ (1) –Є –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ (2) —А–∞—Б—В–µ–љ–Є–є
—А–µ–і–Є—Б–∞ –Њ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ –Є–Ј—Г—З–µ–љ–Є—О –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–Њ—Б—В–∞ –Є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ—А–Є —А–∞–Ј–ї–Є—З–љ—Л—Е (–≤–њ–ї–Њ—В—М –і–Њ –љ–∞—Б—Л—Й–∞—О—Й–Є—Е) –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞, –њ–Њ–Ј–≤–Њ–ї–Є–ї–Є —Г—Б—В–∞–љ–Њ–≤–Є—В—М, —З—В–Њ —Б–≤–µ—В–Њ–≤–Њ–µ –љ–∞—Б—Л—Й–µ–љ–Є–µ –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ —А–Њ—Б—В–∞ –љ–∞—Б—В—Г–њ–∞–µ—В –њ—А–Є –±–Њ–ї–µ–µ –Є–Є–Ј–Ї–Є—Е —Г—А–Њ–≤–љ—П—Е –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є, —З–µ–Љ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј [5, 6].
–Т –Ї–∞—З–µ—Б—В–≤–µ –Є–ї–ї—О—Б—В—А–∞—Ж–Є–Є –њ—А–Є–≤–Њ–і—П—В—Б—П —Д–Њ—В–Њ–≥—А–∞—Д–Є–Є —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ—А–Є –љ–Є–Ј–Ї–Є—Е, –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л—Е –Є –љ–∞—Б—Л—Й–∞—О—Й–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е –§–Р–†. –Э–∞ —А–Є—Б. 3 –њ–Њ–Ї–∞–Ј–∞–љ –≤–љ–µ—И–љ–Є–є –≤–Є–і —А–∞—Б—В–µ–љ–Є–є —Б–∞–ї–∞—В–∞ –≤ –≤–Њ–Ј—А–∞—Б—В–µ 20 –і–љ–µ–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ–Њ–і —Б–≤–µ—В–Њ–Љ –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л—Е –ї–∞–Љ–њ –њ—А–Є —А–∞–Ј–љ—Л—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞. –Т–Є–і–љ–Њ, —З—В–Њ –і–ї—П —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ—А–Є –љ–Є–Ј–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞, —Е–∞—А–∞–Ї—В–µ—А–љ—Л –і–ї–Є–љ–љ—Л–µ —З–µ—А–µ—И–Ї–Є, —В–Њ–љ–Ї–Є–µ –≤—Л—В—П–љ—Г—В—Л–µ –ї–Є—Б—В—М—П. –Я—А–Є –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ (–Њ–Ї–Њ–ї–Њ 200 –Т—В/–Љ2 –§–Р–†) —А–∞—Б—В–µ–љ–Є—П –Є–Љ–µ–ї–Є –љ–µ —В–Њ–ї—М–Ї–Њ –і–Њ—Б—В–∞—В–Њ—З–љ—Г—О –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤, –љ–Њ –Є –≤—Л—Б–Њ–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј —Б –µ–і–Є–љ–Є—Ж—Л –µ–µ –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є, —В. —Б. —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є —А–Њ—Б—В –±—Л–ї–Є —Е–Њ—А–Њ—И–Њ —Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ—Л.
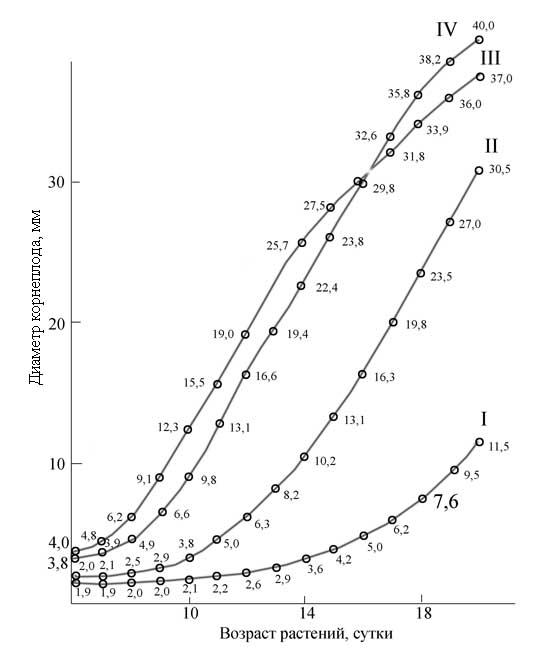
–†–Є—Б. 5. –°—Г—В–Њ—З–љ—Л–є –њ—А–Є—А–Њ—Б—В –і–Є–∞–Љ–µ—В—А–∞ –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ —Г —А–∞—Б—В–µ–љ–Є–є —А–µ–і–Є—Б–∞ –Њ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞ (–Ї—Б–µ–љ–Њ–љ–Њ–≤—Л–µ –ї–∞–Љ–њ—Л) I - 50, II - 140, III - 325, IV - 410 –Т—В/–Љ2
–Э–∞ —А–Є—Б. 4 –њ–Њ–Ї–∞–Ј–∞–љ–∞ –Ј–∞–≤–Є—Б–Є–Љ–Њ—Б—В—М —А–Њ—Б—В–∞ –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ –Є –ї–Є—Б—В—М–µ–≤ —А–µ–і–Є—Б–∞ –Њ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–†. –Ь–Њ–ґ–љ–Њ –≤–Є–і–µ—В—М, —З—В–Њ –њ–Њ –Љ–µ—А–µ —Г–≤–µ–ї–Є—З–µ–љ–Є—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞ –≤–µ—Б –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ —А–∞—Б—В–µ—В, –∞ –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤ —Г–Љ–µ–љ—М—И–∞–µ—В—Б—П.
–С—Л–ї–Є –њ—А–Њ–≤–µ–і–µ–љ—Л —В–∞–Ї–ґ–µ –Њ–њ—Л—В—Л, –≤ –Ї–Њ—В–Њ—А—Л—Е –µ–ґ–µ–і–љ–µ–≤–љ–Њ –Є–Ј–Љ–µ—А—П–ї—Б—П –і–Є–∞–Љ–µ—В—А –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ —Г —А–∞—Б—В–µ–љ–Є–є —А–µ–і–Є—Б–∞, —А–∞—Б—В—Г—Й–Є—Е –њ—А–Є –≤—Л—Б–Њ–Ї–Є—Е –Є –љ–Є–Ј–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л—Е –ї–∞–Љ–њ. –Ь–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л–є –њ—А–Є—А–Њ—Б—В –і–Є–∞–Љ–µ—В—А–∞ –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ –љ–∞–±–ї—О–і–∞–ї—Б—П –њ—А–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–† 325 –Т—В/–Љ2 (—А–Є—Б. 5).
–°–ї–µ–і—Г–µ—В –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ –њ—А–Є –≤—Л—Б–Њ–Ї–Њ–Љ —Г—А–Њ–≤–љ–µ –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є –Є–љ—В–µ–љ—Б–Є–≤–љ—Л–є —А–Њ—Б—В –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ –љ–∞—З–Є–љ–∞–ї—Б—П —Г–ґ–µ —Б 6-–і–љ–µ–≤–љ–Њ–≥–Њ –≤–Њ–Ј—А–∞—Б—В–∞, –≤ —В–Њ –≤—А–µ–Љ—П –Ї–∞–Ї –њ—А–Є –љ–Є–Ј–Ї–Њ–є –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є –Ј–∞–Љ–µ—В–љ—Л–є –њ—А–Є—А–Њ—Б—В –љ–∞–±–ї—О–і–∞–ї—Б—П —В–Њ–ї—М–Ї–Њ —Б 14-–і–љ–µ–≤–љ–Њ–≥–Њ –≤–Њ–Ј—А–∞—Б—В–∞. –Ю–і–љ–∞–Ї–Њ –њ—А–Є –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Б–≤–µ—В–∞ (410 –Т—В/–Љ2 –§–Р–†) —Г —А–∞—Б—В–µ–љ–Є–є —Б 16-–і–љ–µ–≤–љ–Њ–≥–Њ –≤–Њ–Ј—А–∞—Б—В–∞ –љ–∞—З–Є–љ–∞–ї–Є –ґ–µ–ї—В–µ—В—М –Є –Њ—В–Љ–Є—А–∞—В—М –љ–Є–ґ–љ–Є–µ –ї–Є—Б—В—М—П, –∞ –Ї 20 –і–љ—О –њ—А–Є—А–Њ—Б—В –Ї–Њ—А–љ–µ–њ–ї–Њ–і–∞ –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –њ—А–µ–Ї—А–∞—Й–∞–ї—Б—П - –љ–∞—Б—В—Г–њ–∞–ї–Њ –Њ—З–µ–љ—М —А–∞–љ–љ–µ–µ —Б—В–∞—А–µ–љ–Є–µ —А–∞—Б—В–µ–љ–Є–є. –Я—А–Є —Б–Є–ї—М–љ–Њ–Љ –Ј–∞–≥—Г—Й–µ–љ–Є–Є –њ–Њ—Б–µ–≤–∞ —Г—А–Њ–≤–µ–љ—М —Б–≤–µ—В–Њ–≤–Њ–≥–Њ –љ–∞—Б—Л—Й–µ–љ–Є—П –њ–Њ —А–Њ—Б—В—Г –Є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј—Г –Љ–Њ–ґ–љ–Њ –њ–Њ–і–љ—П—В—М [9]. –Т —Н—В–Њ–Љ —Б–ї—Г—З–∞–µ –≤–µ—А—Е–љ–Є–µ –ї–Є—Б—В—М—П —Б–ї—Г–ґ–∞—В –Ј–∞—Й–Є—В–љ—Л–Љ —Н–Ї—А–∞–љ–Њ–Љ –Њ—В —З—А–µ–Ј–Љ–µ—А–љ–Њ –≤—Л—Б–Њ–Ї–Њ–≥–Њ —Г—А–Њ–≤–љ—П –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є. –Т –≤–∞—А–Є–∞–љ—В–∞—Е —Б –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М—О –§–Р–† 320 –Є 410 –Т—В/–Љ2 –≤—Л—Б–Њ–Ї–∞—П –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М- —А–∞—Б—В–µ–љ–Є–є –±—Л–ї–∞ –њ–Њ–ї—Г—З–µ–љ–∞ —В–Њ–ї—М–Ї–Њ –њ—А–Є —Г—Б–ї–Њ–≤–Є–Є –њ—А–Є–Љ–µ–љ–µ–љ–Є—П –њ–Њ–≤—Л—И–µ–љ–љ—Л—Е –і–Њ–Ј –∞–Ј–Њ—В–∞ –Є –Љ–∞–≥–љ–Є—П (—В—А–Є –љ–Њ—А–Љ—Л –њ–Њ –Ъ–љ–Њ–њ—Г). –Я–Њ–і–Ї–Њ—А–Љ–Ї–∞ —А–∞—Б—В–µ–љ–Є–є –≤ –њ–Њ—З–≤–µ–љ–љ–Њ–є –Ї—Г–ї—М—В—Г—А–µ –њ—А–Њ–≤–Њ–і–Є–ї–∞—Б—М 2 —А–∞–Ј–∞ –≤ –љ–µ–і–µ–ї—О.
–Э–∞ —А–Є—Б. 6 –њ–Њ–Ї–∞–Ј–∞–љ—Л —А–∞—Б—В–µ–љ–Є—П –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞ —Б–Њ—А—В–∞ –Я–µ—А–µ–і–Њ–≤–Є–Ї (–≤—Л—Б–Њ–Ї–Њ—А–Њ—Б–ї—Л–є —Б–Њ—А—В), –≤—Л—А–∞—Й–µ–љ–љ—Л–µ –њ—А–Є –і–≤—Г—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е –§–Р–†: 250-300 –Є 130 –Т—В/–Љ2. –†–∞—Б—В–µ–љ–Є—П –≤–∞—А–Є–∞–љ—В–∞ 250300 –Т—В/–Љ2 –Є–Љ–µ–ї–Є –≤ 2 —А–∞–Ј–∞ –Љ–µ–љ—М—И—Г—О –≤—Л—Б–Њ—В—Г —Б—В–µ–±–ї—П, –Љ–µ–љ—М—И—Г—О –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤, –љ–Њ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј (—В–∞–±–ї. 1). –†–∞—Б—З–µ—В—Л –њ–Њ —Б—Г—В–Њ—З–љ–Њ–Љ—Г —Г—Б–≤–Њ–µ–љ–Є—О –°–Ю2 –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –љ–µ–±–Њ–ї—М—И–∞—П –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤ —Г —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞ –љ–∞ –≤—Л—Б–Њ–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е –§–Р–† –Ї–Њ–Љ–њ–µ–љ—Б–Є—А–Њ–≤–∞–ї–∞—Б—М –≤—Л—Б–Њ–Ї–Є–Љ —Г—А–Њ–≤–љ–µ–Љ –∞—Б—Б–Є–Љ–Є–ї—П—Ж–Є–Є –°–Ю2 .

–†–Є—Б. 6. –†–µ–∞–Ї—Ж–Є—П –Є–љ–≥–Є–±–Є—А–Њ–≤–∞–љ–Є–є —А–Њ—Б—В–∞ —Б—В–µ–±–ї—П –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞ (—Б–Њ—А—В –Я–µ—А–µ–і–Њ–≤–Є–Ї) —Б–≤–µ—В–Њ–Љ –≤—Л—Б–Њ–Ї–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є (–Т—В/–Љ2 –§–Р–†) 1 - 250-300, 2 - 130.
–Т —Н—В–Њ–Љ –Њ–њ—Л—В–µ –≤ –Њ–±–Њ–Є—Е –≤–∞—А–Є–∞–љ—В–∞—Е —А–∞—Б–њ—А–µ–і–µ–ї–µ–љ–Є–µ –∞—Б—Б–Є–Љ–Є–ї—П—В–Њ–≤ –њ–Њ –Њ—А–≥–∞–љ–∞–Љ –±—Л–ї–Њ —А–∞–Ј–ї–Є—З–љ—Л–Љ: –њ—А–Є –љ–Є–Ј–Ї–Њ–є –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є –Њ—Б–љ–Њ–≤–љ–∞—П –Љ–∞—Б—Б–∞ –∞—Б—Б–Є–Љ–Є–ї—П—В–Њ–≤ –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–∞—Б—М –љ–∞ —А–Њ—Б—В –≤–µ–≥–µ—В–∞—В–Є–≤–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤ - –ї–Є—Б—В—М–µ–≤, —Б—В–µ–±–ї—П, –∞ –њ—А–Є –≤—Л—Б–Њ–Ї–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–† –∞—Б—Б–Є–Љ–Є–ї—П—В—Л –≤ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ–є —Б—В–µ–њ–µ–љ–Є –Є—Б–њ–Њ–ї—М–Ј–Њ–≤–∞–ї–Є—Б—М –љ–∞ —А–Њ—Б—В —А–µ–њ—А–Њ–і—Г–Ї—В–Є–≤–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤ - –Ї–Њ—А–Ј–Є–љ–Ї–Є —Б –±–Њ–ї—М—И–Є–Љ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ–Љ –≤ –љ–µ–є –≤—Л–њ–Њ–ї–љ–µ–љ–љ—Л—Е —Б–µ–Љ—П–љ–Њ–Ї (—А–Є—Б. 6). –Ф–ї—П —Г–і–Њ–±—Б—В–≤–∞ –Њ—Ж–µ–љ–Ї–Є –Є —Б—В–∞–љ–і–∞—А—В–Є–Ј–∞—Ж–Є–Є –њ–Њ–ї—Г—З–∞–µ–Љ—Л—Е —А–µ–Ј—Г–ї—М—В–∞—В–Њ–≤ –і–ї—П —Г—Б–ї–Њ–≤–Є–є —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л –љ–∞ –Њ—Б–љ–Њ–≤–∞–љ–Є–Є –љ–∞—И–Є—Е —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л—Е –і–∞–љ–љ—Л—Е –Љ–Њ–ґ–љ–Њ –і–∞—В—М —Б–ї–µ–і—Г—О—Й–Є–µ –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П —В—А–µ–Љ –Њ—Б–љ–Њ–≤–љ—Л–Љ –і–Є–∞–њ–∞–Ј–Њ–љ–∞–Љ –њ—А–Є–Љ–µ–љ—П–µ–Љ—Л—Е –Є–љ—В–µ–Є—Б–Є–≤–љ–Њ—Б—В–µ–є –§–Р–†.
- –Э–Є–Ј–Ї–∞—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М –§–Р–† (–Љ–Є–љ–Є–Љ–∞–ї—М–љ–Њ –і–Њ–њ—Г—Б—В–Є–Љ–∞—П) - –і–Є–∞–њ–∞–Ј–Њ–љ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є, –њ—А–Є –Ї–Њ—В–Њ—А–Њ–Љ —А–Њ—Б—В –≤–µ–≥–µ—В–∞—В–Є–≤–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В, –љ–Њ –љ–µ –Њ–±—А–∞–Ј—Г—О—В—Б—П –њ–Њ–ї–љ–Њ—Ж–µ–љ–љ—Л–µ –≥–µ–љ–µ—А–∞—В–Є–≤–љ—Л–µ –Њ—А–≥–∞–љ—Л. –Я—А–Є —Н—В–Њ–Љ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є–Љ–µ–µ—В –љ–Є–Ј–Ї–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є. –Ф–ї—П —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є —Н—В–Њ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –≤ –і–Є–∞–њ–∞–Ј–Њ–љ–µ 15-30 –Т—В/–Љ2.
- –Ю–њ—В–Є–Љ–∞–ї—М–љ–∞—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М –§–Р –† - –і–Є–∞–њ–∞–Ј–Њ–љ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є, –њ—А–Є –Ї–Њ—В–Њ—А–Њ–Љ –љ–∞–±–ї—О–і–∞–µ—В—Б—П –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–µ –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–µ –±–Є–Њ–Љ–∞—Б—Б—Л –≤ –µ–і–Є–љ–Є—Ж—Г –≤—А–µ–Љ–µ–љ–Є. –Я—А–Є —Н—В–Њ–Љ –Ъ—Е–Њ–Ј —Б–Њ—Б—В–∞–≤–ї—П–µ—В 50-70%. –§–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є —А–Њ—Б—В —Е–Њ—А–Њ—И–Њ —Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ—Л. –Ф–ї—П —Б–≤–µ—В–Њ–ї—О–±–Є–≤—Л—Е —А–∞—Б—В–µ–љ–Є–є —Н—В–Њ –і–Є–∞–њ–∞–Ј–Њ–љ –§–Р–† 150-220 –Т—В/–Љ2.
–Э–∞—Б—Л—Й–∞—О—Й–∞—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М –§–Р–† - –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–∞—П –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М —Б–≤–µ—В–∞, –њ—А–Є –Ї–Њ—В–Њ—А–Њ–є –і–Њ—Б—В–Є–≥–∞–µ—В—Б—П –≤—Л—Е–Њ–і —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –љ–∞ –њ–ї–∞—В–Њ —Б–≤–µ—В–Њ–≤–Њ–≥–Њ –љ–∞—Б—Л—Й–µ–љ–Є—П, —В. –µ. –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј. –Я—А–Є —Н—В–Њ–Љ, –љ–∞–±–ї—О–і–∞–µ—В—Б—П —В–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ —А–Њ—Б—В–∞ —Б—В–µ–±–ї—П –Є –і—А—Г–≥–Є—Е –Њ—Б–µ–≤—Л—Е –Њ—А–≥–∞–љ–Њ–≤ - —А–∞—Б—В–µ–љ–Є—П –њ—А–Є–Њ–±—А–µ—В–∞—О—В –љ–Є–Ј–Ї–Њ—А–Њ—Б–ї—Г—О —Д–Њ—А–Љ—Г. –Ъ—Е–Њ–Ј –≤—Л—Б–Њ–Ї–Є–є, –љ–Њ –Њ–±—Й–µ–є –±–Є–Њ–Љ–∞—Б—Б—Л –љ–∞–Ї–∞–њ–ї–Є–≤–∞–µ—В—Б—П –Љ–µ–љ—М—И–µ, —З–µ–Љ –њ—А–Є –Њ–њ—В–Є–Љ–∞–ї—М–љ–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є, —В–∞–Ї –Ї–∞–Ї —А–Њ—Б—В –Ј–∞—В–Њ—А–Љ–Њ–ґ–µ–љ (400 –Т—В –Є –±–Њ–ї–µ–µ)
|
–Я–Њ–Ї–∞–Ј–∞—В–µ–ї–Є
|
–Ш–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М —Б–≤–µ—В–∞ –Т—В/–Љ.–Ї–≤ –§–Р–† |
|
|
250 - 300 |
130 |
|
| –Т—Л—Б–Њ—В–∞ —А–∞—Б—В–µ–љ–Є–є —Б–Љ | 74 ± 2,730 | 154,6 ± 3,07 |
| –Ш–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –Љ–≥ –°–Ю2 /–і–Љ *—З | 56 ± 2,047 | 25,5 ±1,682 |
| –І–Є—Б–ї–Њ –ї–Є—Б—В—М–µ–≤ | 24,5 ± 0,5 | 25,6 ± 0,3 |
| –Я–ї–Њ—В–љ–Њ—Б—В—М –ї–Є—Б—В—М–µ–≤, –і–Љ2 | 52,2 ± 1,365 | 121,6 ± 3,07 |
| –Ф–Є–∞–Љ–µ—В—А –Ї–Њ—А–Ј–Є–љ–Ї–Є, —Б–Љ | 25,6 ± 0,443 | 14,8–Љ ± 0,0512 |
| –Ь–∞—Б—Б–∞ –≤—Л–њ–Њ–ї–љ–µ–љ–љ—Л—Е —Б–µ–Љ—П–љ–Њ–Ї –љ–∞ —А–∞—Б—В–µ–љ–Є–µ, –≥ | 86,3 ± 3,024 | 61,2 ± 1,365 |
| –°—Г—Е–∞—П –±–Є–Њ–Љ–∞—Б—Б–∞ —А–∞—Б—В–µ–љ–Є—П, –≥ | 383,6 ± 6,731 | 565,3 ± 9,0706 |
–Ґ–∞–±–ї–Є—Ж–∞ 1. –•–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–∞ —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞ –≤ –≤–Њ–Ј—А–∞—Б—В–µ 60 –і–љ–µ–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ—А–Є —А–∞–Ј–љ—Л—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л—Е –ї–∞–Љ–њ.
–Ш–Ј–≤–µ—Б—В–љ–Њ, —З—В–Њ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј —В–∞–Ї –ґ–µ, –Ї–∞–Ї –Є —А–Њ—Б—В, –љ–∞—Е–Њ–і–Є—В—Б—П –њ–Њ–і –≥–Њ—А–Љ–Њ–љ–∞–ї—М–љ–Њ–Є–љ–≥–Є–±–Є—В–Њ—А–љ—Л–Љ –Ї–Њ–љ—В—А–Њ–ї–µ–Љ [10], –∞ —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–∞ —П–≤–ї—П—О—В—Б—П –Љ–µ—Б—В–Њ–Љ —Б–Є–љ—В–µ–Ј–∞ –љ–µ —В–Њ–ї—М–Ї–Њ –љ–µ–Ї–Њ—В–Њ—А—Л—Е —Д–µ–љ–Њ–ї–Њ–≤, –љ–Њ —В–∞–Ї–ґ–µ –≥–Є–±–±–µ—А–µ–ї–ї–Є–љ–Њ–≤ –Є –∞–±—Б—Ж–Є–Ј–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л [11]. –Т —Б–≤–Њ—О –Њ—З–µ—А–µ–і—М –±–Є–Њ—Б–Є–љ—В–µ–Ј —Н—В–Є—Е –≤–µ—Й–µ—Б—В–≤ –љ–∞—Е–Њ–і–Є—В—Б—П –њ–Њ–і –њ—А—П–Љ—Л–Љ –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–Љ –Ї–Њ–љ—В—А–Њ–ї–µ–Љ, —Б –Њ–і–љ–Њ–є —Б—В–Њ—А–Њ–љ—Л, –Є –њ–Њ–і –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–µ–Љ –≤–љ–µ—И–љ–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤ (—Б–≤–µ—В, —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–µ —Н–ї–µ–Љ–µ–љ—В–Њ–≤ –Љ–Є–љ–µ—А–∞–ї—М–љ–Њ–≥–Њ –њ–Є—В–∞–љ–Є—П) -—Б –і—А—Г–≥–Њ–є.
–†–∞–љ–µ–µ –љ–∞–Љ–Є –±—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ —Б–≤–µ—В –≤—Л—Б–Њ–Ї–Є—Е (–љ–∞—Б—Л—Й–∞—О—Й–Є—Е) –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–µ–є –≤—Л–Ј—Л–≤–∞–µ—В —Б–љ–Є–ґ–µ–љ–Є–µ —Б–Њ–і–µ—А–ґ–∞–љ–Є—П –≤ —А–∞—Б—В–µ–љ–Є—П—Е —Д–Є—В–Њ–≥–Њ—А–Љ–Њ–љ–Њ–≤ –Є –љ–∞–Ї–Њ–њ–ї–µ–љ–Є–µ —Д—А–∞–≤–Њ–љ–Њ–Є–і–љ—Л—Е —Б–Њ–µ–і–Є–љ–µ–љ–Є–є –Є –њ—А–Є—А–Њ–і–љ—Л—Е –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–≤ —А–Њ—Б—В–∞ [5, 6]. –Я—А–Є —Н—В–Њ–Љ –Љ–µ–љ—П–µ—В—Б—П —Б–Ї–Њ—А–Њ—Б—В—М —А–Њ—Б—В–∞ –Є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–є. –Я—А–Є –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–µ –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–≤ —А–Њ—Б—В–∞ –Є —Д–Є—В–Њ–≥–Њ—А–Љ–Њ–љ–Њ–≤ —Е–Њ—А–Њ—И–Њ —Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ–Њ - –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –Њ–њ—В–Є–Љ–∞–ї—М–љ–∞—П —Б–∞–Љ–Њ—А–µ–≥—Г–ї—П—Ж–Є—П —А–Њ—Б—В–Њ–≤—Л—Е –њ—А–Њ—Ж–µ—Б—Б–Њ–≤.
–Ф–ї–Є—В–µ–ї—М–љ–Њ–µ –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–µ —А–∞—Б—В–µ–љ–Є–є –њ—А–Є –≤—Л—Б–Њ–Ї–Є—Е –Є–ї–Є –љ–Є–Ј–Ї–Є—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е —Б–≤–µ—В–∞ –Њ–±–µ—Б–њ–µ—З–Є–≤–∞–µ—В —А–∞–Ј–≤–Є—В–Є–µ –Ї–Њ–Љ–њ–µ–љ—Б–∞—В–Њ—А–љ—Л—Е –Љ–µ—Е–∞–љ–Є–Ј–Љ–Њ–≤, –љ–Њ—Б—П—Й–Є—Е –∞–і–∞–њ—В–Є–≤–љ—Л–є —Е–∞—А–∞–Ї—В–µ—А. –Ґ–∞–Ї, –≤—Л—Б–Њ–Ї–Є–µ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–† –≤—Л–Ј—Л–≤–∞—О—В —А–µ–Ј–Ї–Њ–µ —В–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ —А–Њ—Б—В–∞ –Њ—Б–µ–≤—Л—Е –Њ—А–≥–∞–љ–Њ–≤, –њ—А–Є —Н—В–Њ–Љ –≤–Ї–ї—О—З–∞—О—В—Б—П —Б–њ–µ—Ж–Є—Д–Є—З–µ—Б–Ї–Є–µ –Є–Є–≥–Є–±–Є—В–Њ—А–љ—Л–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л, –њ—А–Њ—П–≤–ї–µ–љ–Є–µ –Ї–Њ—В–Њ—А—Л—Е –љ–∞ —Б–ї–∞–±—Л—Е –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—П—Е –љ–µ –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М [5, 6, 12]. –Ґ–∞–Ї–Њ–≤—Л –Ј–∞–Ї–Њ–љ–Њ–Љ–µ—А–љ–Њ—Б—В–Є, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –і–ї—П —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–µ –Ї–Њ—В–Њ—А—Л—Е –њ—А–Њ–≤–Њ–і–Є–ї–Њ—Б—М –≤ —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А–µ –њ–Њ–і –±–µ–ї—Л–Љ —Б–≤–µ—В–Њ–Љ –њ–Њ —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–є —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–µ –≤ –Њ–±–ї–∞—Б—В–Є –§–Р–†, –±–ї–Є–Ј–Ї–Њ–є –Ї —Б–Њ–ї–љ–µ—З–љ–Њ–Љ—Г —Б–њ–µ–Ї—В—А—Г (–Ї—Б–µ–љ–Њ–љ–Њ–≤—Л–µ –ї–∞–Љ–њ—Л).
–Ъ–∞–Ї–Њ–≤–∞ –ґ–µ –Њ—В–≤–µ—В–љ–∞—П —А–µ–∞–Ї—Ж–Є—П —А–∞—Б—В–µ–љ–Є–є –љ–∞ –Њ–±–ї—Г—З–µ–љ–Є–µ –Є—Е —В—А–µ–Љ—П –Њ—Б–љ–Њ–≤–љ—Л–Љ–Є –Њ–±–ї–∞—Б—В—П–Љ–Є –§–Р–† - —Б–Є–љ–µ–є, –Ї—А–∞—Б–љ–Њ–є –Є–ї–Є –Ј–µ–ї–µ–љ–Њ–є? –≠—В–Њ—В –≤–Њ–њ—А–Њ—Б –≤–∞–ґ–µ–љ –і–ї—П —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л —А–∞—Б—В–µ–љ–Є–є –≤ —Б–≤—П–Ј–Є —Б —А–∞–Ј—А–∞–±–Њ—В–Ї–Њ–є –љ–Њ–≤—Л—Е —А–∞—Б—В–µ–љ–Є–µ–≤–Њ–і—З–µ—Б–Ї–Є—Е –Є—Б—В–Њ—З–љ–Є–Ї–Њ–≤ —Б–≤–µ—В–∞, –Њ–љ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В —В–∞–Ї–ґ–µ –Є —В–µ–Њ—А–µ—В–Є—З–µ—Б–Ї–Є–є –Є–љ—В–µ—А–µ—Б, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Г –≤—Б–µ—Е –≤—Л—Б—И–Є—Е —А–∞—Б—В–µ–љ–Є–є –Є–Љ–µ—О—В—Б—П —А–∞–Ј–ї–Є—З–љ—Л–µ —Д–Њ—В–Њ—А–µ—Ж–µ–њ—В–Њ—А—Л, –Њ–±–µ—Б–њ–µ—З–Є–≤–∞—О—Й–Є–µ –њ–Њ–≥–ї–Њ—Й–µ–љ–Є–µ —Н–љ–µ—А–≥–Є–Є –њ–Њ –≤—Б–µ–є –Њ–±–ї–∞—Б—В–Є –§–Р–†.
–Т –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П –і–ї—П —Ж–µ–ї–µ–є —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л –≤—Л–њ—Г—Б–Ї–∞—О—В —А–∞–Ј–ї–Є—З–љ—Л–µ —В–Є–њ—Л –Ь–У–Ы, –Ї–Њ—В–Њ—А—Л–µ, –Ї —Б–Њ–ґ–∞–ї–µ–љ–Є—О, –Є–Љ–µ—О—В –љ–µ —Б–њ–ї–Њ—И–љ–Њ–є, –∞ –ї–Є–љ–µ–є—З–∞—В—Л–є —Б–њ–µ–Ї—В—А –Є –љ–µ–і–Њ—Б—В–∞—В–Њ—З–љ–Њ–µ –Є–Ј–ї—Г—З–µ–љ–Є–µ (–Ї—А–Њ–Љ–µ –ї–∞–Љ–њ—Л –Ф–†–Ю–Ґ) –≤ –Ї—А–∞—Б–љ–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞ 660-680 –љ–Љ. –Ґ–µ–Љ –љ–µ –Љ–µ–љ–µ–µ –Ь–У–Ы —П–≤–ї—П—О—В—Б—П –љ–∞–Є–±–Њ–ї–µ–µ –њ–µ—А—Б–њ–µ–Ї—В–Є–≤–љ—Л–Љ–Є, —В–∞–Ї –Ї–∞–Ї –Є–Љ–µ—О—В –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–є –Ъ–Я–Ф (~30%) –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б –Ї—Б–µ–љ–Њ–љ–Њ–≤—Л–Љ–Є –ї–∞–Љ–њ–∞–Љ–Є (–Ъ–Я–Ф –Ї–Њ—В–Њ—А—Л—Е —А–∞–≤–µ–љ 12%).
| –¶–≤–µ—В–љ—Л–µ –ї—О–Љ–Є–љ–Є—Б—Ж–µ–љ —В–љ—Л–µ –ї–∞–Љ–њ—Л |
–°—Л—А–∞—П –±–Є–Њ–Љ–∞—Б—Б–∞ |
–Ю–±—Й–∞—П —Б—Л—А–∞—П –±–Є–Њ–Љ–∞—Б—Б–∞ –Њ–і–љ–Њ–≥–Њ —А–∞—Б—В–µ–љ–Є –≥. | –•–ї (a+b), –Љ–≥*–і–Љ-2 | –Я–Њ–≥–ї–Њ—Й–µ–љ–Є–µ –°–Ю2 –Љ–≥*–і–Љ-2*—З-1 | |||
|
–Ы–Є—Б—В—М—П |
–°—В–µ–±–ї–Є –Є —З–µ—А–µ—И–Ї |
–Ъ–Њ—А–љ–Є |
–Я–ї–Њ–і—Л |
||||
| –°–Є–љ–µ–≥–Њ —Б–≤–µ—В–∞ –Ы–°-150 | 31,66±2,38 | 29,33±1,706 | 10,02±0,682 | 133,3±4,095 | 204,33±3,341 | 8,078±0,31 | 24,5±1,36 |
| –Ъ—А–∞—Б–љ–Њ–≥–Њ —Б–≤–µ—В–∞ –Ы–Ъ-65 | 55,0±2,68 | 42,16±1,365 | 13,66±1,023 | 186,1±3,071 | 296,83±4,436 | 5,6±0,09 | 20,3±1,25 |
| –С–µ–ї–Њ–≥–Њ —Б–≤–µ—В–∞ –Ы–•–С-150 | 39,66±3,07 | 31,66±1,706 | 8,3±0,375 | 99,33±6,143 | 178,95±8,156 | 5,01±0,06 | 18,2±1,45 |
| –Ы–§–†-150 | 48,0±1,36 | 40,66±1,365 | 18,0±0,682 | 216,0±5,119 | 322,6±4,778 | 5,78±0,2 | 22,4±1,01 |
| –С–µ–ї–Њ–≥–Њ —Б–≤–µ—В–∞ –Ы–•–С-150 | 42,2±3,87 | 34,4±1,91 | 12,1±1,22 | 156,7±4,88 | 245,4±6,68 | 5,9±0,22 | 21,7±1,2 |
–Ґ–∞–±–ї–Є—Ж–∞ 2. –•–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–∞ —А–∞—Б—В–µ–љ–Є–є –њ–µ—А—Ж–∞ —Б–Њ—А—В–∞ –Т–Є–љ–Є-–њ—Г—Е, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ–Њ–і –Є–Ј–ї—Г—З–µ–љ–Є–µ–Љ —Ж–≤–µ—В–љ—Л—Е –ї—О–Љ–Є–љ–Є—Б—Ж–µ–љ—В–љ—Л—Е –ї–∞–Љ–њ.
–§–Р–† –≤—Л—А–∞–≤–љ–µ–љ–∞ –њ–Њ —З–Є—Б–ї—Г –њ–∞–і–∞—О—Й–Є—Е –Ї–≤–∞–љ—В–Њ–≤ –Є —А–∞–≤–љ–∞ 19,7*1015 –Ї–≤–∞–љ—В*—Б–Љ-2*—Б-1, –≤–Њ–Ј—А–∞—Б—В —А–∞—Б—В–µ–љ–Є–є 80 –і–љ–µ–є.
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є—П –њ–Њ –Њ—Ж–µ–љ–Ї–µ –і–µ–є—Б—В–≤–Є—П —Б–≤–µ—В–∞ –њ—А—П–Љ–Њ–≥–Њ —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –љ–∞ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј, —А–Њ—Б—В –Є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–є –Љ—Л –њ—А–Њ–≤–Њ–і–Є–ї–Є –њ—А–Є –і–ї–Є—В–µ–ї—М–љ–Њ–Љ –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–Є —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і –Є—Б—В–Њ—З–љ–Є–Ї–∞–Љ–Є —Б–≤–µ—В–∞, –Є–Љ–µ—О—Й–Є–Љ–Є –Љ–∞–Ї—Б–Є–Љ—Г–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ –Ї—А–∞—Б–љ–Њ–є, —Б–Є–љ–µ–є –Є–ї–Є –Ј–µ–ї–µ–љ–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞. –°–≤–µ—В–Њ–≤—Л–µ –Ї—А–Є–≤—Л–µ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –њ–Њ–ї—Г—З–∞–ї–Є –љ–∞ –±–µ–ї–Њ–Љ —Б–≤–µ—В—Г. –°—Г—Й–µ—Б—В–≤—Г–µ—В –Є –і—А—Г–≥–Њ–є –Љ–µ—В–Њ–і –Њ—Ж–µ–љ–Ї–Є —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ —Б–≤–µ—В–∞ –њ—А–Є –Ї—А–∞—В–Ї–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–Љ –µ–≥–Њ –≤–Њ–Ј–і–µ–є—Б—В–≤–Є–Є, –љ–∞–њ—А–Є–Љ–µ—А, –њ—А–Є –Є–Ј—Г—З–µ–љ–Є–Є —Б–њ–µ–Ї—В—А–Њ–≤ –і–µ–є—Б—В–≤–Є—П —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞, –Ї–Њ–≥–і–∞ —А–∞—Б—В–µ–љ–Є—П –≤—Л—А–∞—Й–Є–≤–∞—О—В –њ–Њ–і –±–µ–ї—Л–Љ —Б–≤–µ—В–Њ–Љ, –∞ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Њ–њ—А–µ–і–µ–ї—П—О—В –≤ —А–∞–Ј–љ—Л—Е —Г—З–∞—Б—В–Ї–∞—Е —Б–њ–µ–Ї—В—А–∞ [13]. –Ю–і–љ–∞–Ї–Њ —Н—В–Њ—В –≤—В–Њ—А–Њ–є –Љ–µ—В–Њ–і –Њ—Ж–µ–љ–Ї–Є –Ї–∞—З–µ—Б—В–≤–∞ —Б–≤–µ—В–∞ –Љ–∞–ї–Њ –њ—А–Є–≥–Њ–і–µ–љ –і–ї—П —Ж–µ–ї–µ–є —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л, –≥–і–µ —Г —А–∞—Б—В–µ–љ–Є–є –њ—А–Є –і–ї–Є—В–µ–ї—М–љ–Њ–є –∞–і–∞–њ—В–∞—Ж–Є–Є –Ї –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–Љ—Г —Б–њ–µ–Ї—В—А—Г –љ–∞–±–ї—О–і–∞—О—В—Б—П –Є–Ј–Љ–µ–љ–µ–љ–Є—П –љ–µ —В–Њ–ї—М–Ї–Њ –≤ —Б–Ї–Њ—А–Њ—Б—В–Є –∞—Б—Б–Є–Љ–Є–ї—П—Ж–Є–Є –°–Ю2 , –љ–Њ –Є –≤ –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Њ–Љ –Є –∞–љ–∞—В–Њ–Љ–Є—З–µ—Б–Ї–Њ–Љ —Б—В—А–Њ–µ–љ–Є–Є —А–∞—Б—В–µ–љ–Є–є [14] –Є –≤ —А–∞–Ј–Љ–µ—А–∞—Е —Г—А–Њ–ґ–∞—П. –Т –Ї–∞—З–µ—Б—В–≤–µ –њ—А–Є–Љ–µ—А–∞ —В–∞–Ї–Њ–є –∞–і–∞–њ—В–∞—Ж–Є–Є –Љ–Њ–ґ–љ–Њ –њ—А–Є–≤–µ—Б—В–Є —А–µ–Ј—Г–ї—М—В–∞—В—Л –Њ–њ—Л—В–Њ–≤ —Б —А–∞—Б—В–µ–љ–Є—П–Љ–Є –њ–µ—А—Ж–∞. –Ы—Г—З—И–Є–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –њ–Њ–і –Є–Ј–ї—Г—З–µ–љ–Є–µ–Љ —Д–Є—В–Њ–ї–∞–Љ–њ –Ы–§–†-150, —Г –Ї–Њ—В–Њ—А—Л—Е –Ї –Ї—А–∞—Б–љ–Њ–Љ—Г —Б–≤–µ—В—Г –±—Л–ї–Њ –і–Њ–±–∞–≤–ї–µ–љ–Њ 25% –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ —Б–Є–љ–µ–є –Њ–±–ї–∞—Б—В–Є (—В–∞–±–ї. 2). –Т —Н—В–Њ–Љ –≤–∞—А–Є–∞–љ—В–µ —А–∞—Б—В–µ–љ–Є—П –±—Л–ї–Є –њ—А–∞–≤–Є–ї—М–љ–Њ —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ—Л –Є –Є–Љ–µ–ї–Є –љ–∞–Є–±–Њ–ї—М—И—Г—О –Љ–∞—Б—Б—Г –њ–ї–Њ–і–Њ–≤. –•–Њ—А–Њ—И–Є–є —А–µ–Ј—Г–ї—М—В–∞—В –±—Л–ї –њ–Њ–ї—Г—З–µ–љ —В–∞–Ї–ґ–µ –њ–Њ–і –Є–Ј–ї—Г—З–µ–љ–Є–µ–Љ –Ї—А–∞—Б–љ—Л—Е –ї–∞–Љ–њ —Б –і–Њ–±–∞–≤–Ї–Њ–є 14% —Б–Є–љ–µ–≥–Њ —Б–≤–µ—В–∞, –≥–і–µ –≤—Б–µ–≥–і–∞ –љ–∞–±–ї—О–і–∞–ї—Б—П –Є–љ—В–µ–љ—Б–Є–≤–љ—Л–є —А–Њ—Б—В –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤, –њ–ї–Њ–і—Л –±—Л–ї–Є —Е–Њ—А–Њ—И–Њ —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ—Л –Є —Б–Њ–Ј—А–µ–≤–∞–ї–Є –љ–∞ –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –і–љ–µ–є —А–∞–љ—М—И–µ, —З–µ–Љ –њ–Њ–і –Ј–µ–ї–µ–љ—Л–Љ –Є —Б–Є–љ–Є–Љ —Б–≤–µ—В–Њ–Љ. –Э–∞–Є–Љ–µ–љ—М—И–Є–є —Г—А–Њ–ґ–∞–є –±—Л–ї –њ–Њ–ї—Г—З–µ–љ –њ–Њ–і –Ј–µ–ї–µ–љ—Л–Љ —Б–≤–µ—В–Њ–Љ. –≠—В–Є —А–∞—Б—В–µ–љ–Є—П –Є–Љ–µ–ї–Є —В–Њ–љ–Ї–Є–µ –ї–Є—Б—В—М—П —Б –љ–Є–Ј–Ї–Њ–є —Г–і–µ–ї—М–љ–Њ–є –Є –Њ–њ—В–Є—З–µ—Б–Ї–Њ–є –њ–ї–Њ—В–љ–Њ—Б—В—М—О, –Љ–µ–љ—М—И–Є–Љ —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ–Љ –•–ї (–∞+–™) –Є —Е—Г–і—И–Є–Љ–Є –њ–Њ–Ї–∞–Ј–∞—В–µ–ї—П–Љ–Є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –≤ –њ–µ—А–µ—Б—З–µ—В–µ –љ–∞. –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞. –£ —А–∞—Б—В–µ–љ–Є–є –љ–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г, –љ–∞–Њ–±–Њ—А–Њ—В, –љ–∞–±–ї—О–і–∞–ї—Б—П –Ј–∞—В–Њ—А–Љ–Њ–ґ–µ–љ–љ—Л–є —А–Њ—Б—В –ї–Є—Б—В—М–µ–≤ –Є –Њ—Б–µ–≤—Л—Е –Њ—А–≥–∞–љ–Њ–≤. –£–і–µ–ї—М–љ–∞—П –њ–ї–Њ—В–љ–Њ—Б—В—М –ї–Є—Б—В—М–µ–≤, —Б–Њ–і–µ—А–ґ–∞–љ–Є–µ –≤ –љ–Є—Е –њ–Є–≥–Љ–µ–љ—В–Њ–≤, –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –Є–Љ–µ–ї–Є –њ—А–Є —Н—В–Њ–Љ –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є, –∞ –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤ –±—Л–ї–∞ –љ–∞–Є–Љ–µ–љ—М—И–µ–є. –Я–Њ –Љ–∞—Б—Б–µ –њ–ї–Њ–і–Њ–≤ —А–∞—Б—В–µ–љ–Є—П –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ —Г—Б—В—Г–њ–∞–ї–Є –≤–∞—А–Є–∞–љ—В—Г –љ–∞ –Ї—А–∞—Б–љ–Њ–Љ —Б–≤–µ—В—Г. –Э–∞ –±–µ–ї–Њ–Љ —Б–≤–µ—В—Г (–ї–∞–Љ–њ—Л –Ы–•–С-150) —А–∞—Б—В–µ–љ–Є—П –Є–Љ–µ–ї–Є –ї—Г—З—И–Є–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є, —З–µ–Љ –њ–Њ–і —Б–Є–љ–Є–Љ –Є –Ј–µ–ї–µ–љ—Л–Љ, –љ–Њ —Е—Г–і—И–Є–µ, —З–µ–Љ –њ–Њ–і –Є–Ј–ї—Г—З–µ–љ–Є–µ–Љ –Ї—А–∞—Б–љ—Л—Е –ї–∞–Љ–њ, —В–∞–Ї –Ї–∞–Ї —Г –±–µ–ї—Л—Е –ї–∞–Љ–њ –Є–Ј–ї—Г—З–µ–љ–Є–µ –≤ –Ї—А–∞—Б–љ–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–Њ—Б—В–∞–≤–ї—П–µ—В –≤—Б–µ–≥–Њ 22-26%.
–Э–∞ –Њ—Б–љ–Њ–≤–∞–љ–Є–Є —Н—В–Є—Е –Њ–њ—Л—В–Њ–≤ –Љ–Њ–ґ–љ–Њ —Б–і–µ–ї–∞—В—М –≤—Л–≤–Њ–і –Њ —В–Њ–Љ, —З—В–Њ –њ–µ—А–µ—Ж —Б–ї–∞–і–Ї–Є–є (–°. –∞–њ–њ–Є–Є—В –ђ.) –Њ—В–љ–Њ—Б–Є—В—Б—П –Ї —З–Є—Б–ї—Г —А–∞—Б—В–µ–љ–Є–є, –і–ї—П –Ї–Њ—В–Њ—А—Л—Е –Љ–∞–Ї—Б–Є–Љ—Г–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П –≤ –Є—Б—В–Њ—З–љ–Є–Ї–µ –і–Њ–ї–ґ–µ–љ –±—Л—В—М –≤ –Ї—А–∞—Б–љ–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞. –°–ї–µ–і—Г–µ—В –Њ—В–Љ–µ—В–Є—В—М, —З—В–Њ —А–∞—Б—В–µ–љ–Є—П –њ–µ—А—Ж–∞, –≤—Л—А–∞—Й–µ–љ–љ—Л–µ –њ—А–Є –љ–µ–≤—Л—Б–Њ–Ї–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –§–Р–†, —Б–Њ–і–µ—А–ґ–∞–ї–Є –љ–∞ 50-60% –±–Њ–ї—М—И–µ –•–ї, —З–µ–Љ –њ—А–Є 200 –Т—В/–Љ2 –§–Р–†. –Ъ–Њ—Н—Д—Д–Є—Ж–Є–µ–љ—В –њ–Њ–≥–ї–Њ—Й–µ–љ–Є—П –§–Р–† —Г —Н—В–Є—Е —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ–Њ–і –ї—О–Љ–Є–љ–µ—Б—Ж–µ–љ—В–љ—Л–Љ–Є –ї–∞–Љ–њ–∞–Љ–Є, —Б–Њ—Б—В–∞–≤–ї—П–ї 90-95%. –Я–µ—А–µ—Ж - –Њ—З–µ–љ—М –њ–ї–∞—Б—В–Є—З–љ–Њ–µ —А–∞—Б—В–µ–љ–Є–µ —Б —Е–Њ—А–Њ—И–µ–є –∞–і–∞–њ—В–∞—Ж–Є–µ–є –Ї —Г—Б–ї–Њ–≤–Є—П–Љ –≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П –Є –±–ї–∞–≥–Њ–і–∞—А—П —Н—В–Њ–Љ—Г –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —В–µ–љ–µ–≤—Л–љ–Њ—Б–ї–Є–≤–Њ. –Ю–і–љ–∞–Ї–Њ –љ–∞–Є–±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–µ —Г—А–Њ–ґ–∞–Є –њ–ї–Њ–і–Њ–≤ –≤ –љ–∞—И–Є—Е –Њ–њ—Л—В–∞—Е –±—Л–ї–Є –њ–Њ–ї—Г—З–µ–љ—Л –њ—А–Є —Г—А–Њ–≤–љ—П—Е –Њ–±–ї—Г—З–µ–љ–љ–Њ—Б—В–Є, —А–∞–≤–љ—Л—Е –њ—А–Є–Љ–µ—А–љ–Њ –њ–Њ–ї–Њ–≤–Є–љ–µ –Њ—В –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л—Е —Б–Њ–ї–љ–µ—З–љ—Л—Е 200- 250 –Т—В/–Љ2 –§–Р–† (–±—Л–ї–Њ –њ–Њ–ї—Г—З–µ–љ–Њ 360-380 –≥ —Б —А–∞—Б—В–µ–љ–Є—П). –Т–Њ–Ј–љ–Є–Ї –≤–Њ–њ—А–Њ—Б, –Ї–∞–Ї–Њ–≤–∞ –њ—А–Є—З–Є–љ–∞ —Б—В–Њ–ї—М –љ–Є–Ј–Ї–Њ–є —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Њ–є –∞–Ї—В–Є–≤–љ–Њ—Б—В–Є –ї–Є—Б—В—М–µ–≤ —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –њ–Њ–і –Ј–µ–ї–µ–љ—Л–Љ —Б–≤–µ—В–Њ–Љ? –Я–Њ—З–µ–Љ—Г —Г —А–∞—Б—В–µ–љ–Є–є, –і–ї–Є—В–µ–ї—М–љ–Њ –∞–і–∞–њ—В–Є—А–Њ–≤–∞–љ–љ—Л—Е –Ї —Б–Є–љ–µ–Љ—Г —Б–≤–µ—В—Г, –љ–∞–±–ї—О–і–∞–ї–Њ—Б—М —Б–∞–Љ–Њ–µ –≤—Л—Б–Њ–Ї–Њ–µ –њ–Њ–≥–ї–Њ—Й–µ–љ–Є–µ –°–Ю2 —Б –µ–і–Є–љ–Є—Ж—Л –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞? –Ф–ї—П –Њ—В–≤–µ—В–∞ –љ–∞ —Н—В–Є –≤–Њ–њ—А–Њ—Б—Л –±—Л–ї–∞ –њ—А–Њ–≤–µ–і–µ–љ–∞ —Б–µ—А–Є—П –Њ–њ—Л—В–Њ–≤ —Б –ї–µ–≤–Ј–µ–µ–є —Б–∞—Д–ї–Њ—А–Њ–≤–Є–і–љ–Њ–є [14]. –Ы–µ–≤–Ј–µ—П - –і–Є–Ї–Њ—А–∞—Б—В—Г—Й–µ–µ —А–∞—Б—В–µ–љ–Є–µ, –Ї–Њ—В–Њ—А–Њ–µ —И–Є—А–Њ–Ї–Њ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Њ –≤ –°–Є–±–Є—А–Є, –Р–ї—В–∞–є—Б–Ї–Њ–Љ –Ї—А–∞–µ, –≤—Б—В—А–µ—З–∞–µ—В—Б—П –Њ–љ–Њ –Є –≤ —Б—А–µ–і–љ–µ–є –њ–Њ–ї–Њ—Б–µ –љ–∞—И–µ–є —Б—В—А–∞–љ—Л. –≠—В–Њ —А–∞—Б—В–µ–љ–Є–µ –Є–љ—В–µ—А–µ—Б–љ–Њ –њ–Њ –і–≤—Г–Љ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї—П–Љ. –Т–Њ-–њ–µ—А–≤—Л—Е, –≤ –Ї–ї–µ—В–Ї–∞—Е —Б—В–Њ–ї–±—З–∞—В–Њ–є –Є –≥—Г–±—З–∞—В–Њ–є –њ–∞—А–µ–љ—Е–Є–Љ—Л –µ–µ –ї–Є—Б—В—М–µ–≤ —Б–Њ–і–µ—А–ґ–Є—В—Б—П –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –љ–µ–±–Њ–ї—М—И–Њ–µ —З–Є—Б–ї–Њ —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–Њ–≤ - 30-40 (—Г –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Ї—Г–ї—М—В—Г—А–љ—Л—Е —А–∞—Б—В–µ–љ–Є–є 100-150), —З—В–Њ –Њ–±–ї–µ–≥—З–∞–µ—В –њ—А–Њ–≤–µ–і–µ–љ–Є–µ –∞–љ–∞–ї–Є–Ј–∞ –≤ —А–∞—Б—З–µ—В–µ –љ–∞ —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В. –Ш –≤–Њ-–≤—В–Њ—А—Л—Е, –Ї–∞–Ї –≤—Б–µ –і–Є–Ї–Њ—А–∞—Б—В—Г—Й–Є–µ —А–∞—Б—В–µ–љ–Є—П, –Њ–љ–Њ –Є–Љ–µ–µ—В –±–Њ–ї–µ–µ —Г—Б—В–Њ–є—З–Є–≤—Г—О –љ–∞—Б–ї–µ–і—Б—В–≤–µ–љ–љ—Г—О –Њ—Б–љ–Њ–≤—Г –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б –Ї—Г–ї—М—В—Г—А–љ—Л–Љ–Є —А–∞—Б—В–µ–љ–Є—П–Љ–Є. –Я–µ—А–µ–і –љ–∞–Љ–Є —Б—В–Њ—П–ї–∞ –Ј–∞–і–∞—З–∞ –≤—Л—П—Б–љ–Є—В—М –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В–Є –∞–і–∞–њ—В–∞—Ж–Є–Є —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ –∞–њ–њ–∞—А–∞—В–∞ –і–Є–Ї–Њ—А–∞—Б—В—Г—Й–µ–≥–Њ —А–∞—Б—В–µ–љ–Є—П –Ї –Ї–∞—З–µ—Б—В–≤—Г —Б–≤–µ—В–∞. –Т —Н—В–Є—Е –Њ–њ—Л—В–∞—Е –±—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –љ–∞ –Ј–µ–ї–µ–љ–Њ–Љ —Б–≤–µ—В—Г —Д–Њ—А–Љ–Є—А—Г–µ—В—Б—П –≤–њ–Њ–ї–љ–µ –∞–Ї—В–Є–≤–љ—Л–є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В, –Њ–љ —В–∞–Ї–ґ–µ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ —Д–Є–Ї—Б–Є—А—Г–µ—В –°–Ю2 , –Ї–∞–Ї –Є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В –љ–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г [14]. –Ю–і–љ–∞–Ї–Њ —З–Є—Б–ї–Њ –Ї–ї–µ—В–Њ–Ї –Є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–Њ–≤ –≤ –µ–і–Є–љ–Є—Ж–µ –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞, —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–љ–Њ–≥–Њ –љ–∞ –Ј–µ–ї–µ–љ–Њ–Љ —Б–≤–µ—В—Г, –±—Л–ї–Њ –љ–∞ 5060% –Љ–µ–љ—М—И–µ, —З–µ–Љ –љ–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г, —З—В–Њ, –≤–µ—А–Њ—П—В–љ–Њ, –Є –њ–Њ—Б–ї—Г–ґ–Є–ї–Њ –њ—А–Є—З–Є–љ–Њ–є –љ–Є–Ј–Ї–Њ–≥–Њ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –љ–∞ –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞.
–Э–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г —З–Є—Б–ї–Њ –Ї–ї–µ—В–Њ–Ї –Є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–Њ–≤ –≤ 1 —Б–Љ2 –ї–Є—Б—В–∞ –±—Л–ї–Њ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –±–Њ–ї—М—И–µ, —З–µ–Љ –љ–∞ –Ї—А–∞—Б–љ–Њ–Љ –Є –Ј–µ–ї–µ–љ–Њ–Љ —Б–≤–µ—В—Г, –Є –љ–∞–±–ї—О–і–∞–ї—Б—П —Б–∞–Љ—Л–є –≤—Л—Б–Њ–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –љ–∞ –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞ [14]. –Ф–ї—П –ї–Є—Б—В—М–µ–≤ —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –љ–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г, –±—Л–ї —Е–∞—А–∞–Ї—В–µ—А–µ–љ –Є –љ–∞–Є–±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–є —Г—А–Њ–≤–µ–љ—М —Б–≤–µ—В–Њ–≤–Њ–≥–Њ –љ–∞—Б—Л—Й–µ–љ–Є—П —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ [15], —З—В–Њ, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –Њ–±—К—П—Б–љ—П–µ—В—Б—П –љ–∞–ї–Є—З–Є–µ–Љ –љ–∞–Є–±–Њ–ї—М—И–µ–≥–Њ —З–Є—Б–ї–∞ —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–Њ–≤, –њ—А–Є—Е–Њ–і—П—Й–Є—Е—Б—П –љ–∞ –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞, –∞ —В–∞–Ї–ґ–µ –Њ—Б–Њ–±–µ–љ–љ–Њ—Б—В—П–Љ–Є –≤–љ—Г—В—А–µ–љ–љ–µ–є –Њ—А–≥–∞–љ–Є–Ј–∞—Ж–Є–Є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–∞ –Є –∞–Ї—В–Є–≤–∞—Ж–Є–µ–є –њ–Њ–і –≤–ї–Є—П–љ–Є–µ–Љ —Б–Є–љ–µ–≥–Њ —Б–≤–µ—В–∞ –±–Є–Њ—Б–Є–љ—В–µ–Ј–∞ —Д–µ—А–Љ–µ–љ—В–Њ–≤ —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Њ–≥–Њ –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ —Г–≥–ї–µ—А–Њ–і–∞ –Є —А–µ–∞–Ї—Ж–Є–є —Ж–µ–њ–Є –њ–µ—А–µ–љ–Њ—Б–∞ —Н–ї–µ–Ї—В—А–Њ–љ–Њ–≤ [16]. –Э–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г –≤ –ї–Є—Б—В—М—П—Е –Њ–±—А–∞–Ј—Г–µ—В—Б—П –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –±–Њ–ї—М—И–µ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–≤ —А–Њ—Б—В–∞ (–∞–±—Б—Ж–Є–Ј–Њ–≤–Њ–є –Ї–Є—Б–ї–Њ—В—Л, –Њ–Ї—Б–Є–Ї–Њ—А–Є—З–љ—Л—Е –Ї–Є—Б–ї–Њ—В –Є –і—А.) –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б —А–∞—Б—В–µ–љ–Є—П–Љ–Є, –≤—Л—А–∞—Й–µ–љ–љ—Л–Љ–Є –љ–∞ –Ї—А–∞—Б–љ–Њ–Љ –Є —В–µ–Љ –±–Њ–ї–µ–µ –љ–∞ –Ј–µ–ї–µ–љ–Њ–Љ —Б–≤–µ—В—Г [6], —З—В–Њ –њ—А–Є–≤–Њ–і–Є—В –Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О —Г–Ї–Њ—А–Њ—З–µ–љ–љ—Л—Е —Б—В–µ–±–ї–µ–є –Є –±–Њ–ї–µ–µ —В–Њ–ї—Б—В—Л—Е –ї–Є—Б—В—М–µ–≤.

–†–Є—Б. 7. –†–∞—Б—В–µ–љ–Є—П –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞ (—Б–Њ—А—В –Я–µ—А–µ–і–Њ–≤–Є–Ї), –≤—Л—А–∞—Й–µ–љ–љ—Л–µ –љ–∞ —Б–Є–љ–µ–Љ (–°) –Є –љ–∞ –Ї—А–∞—Б–љ–Њ–Љ (–Ъ), —Б–Њ–і–µ—А–ґ–∞–≤—И–µ–Љ 14% —Б–Є–љ–µ–≥–Њ (–Ъ), —Б–≤–µ—В—Г. –І–µ—А–µ—И–Ї–Є –ї–Є—Б—В—М–µ–≤ –Є–Ј–Њ–≥–љ—Г—В—Л –Ї–љ–Є–Ј—Г - —А–µ–∞–Ї—Ж–Є—П —Н–њ–Є–љ–∞—Б—В–Є–Є –љ–∞ –Ї—А–∞—Б–љ—Л–є —Б–≤–µ—В.
–Т –Њ–њ—Л—В–∞—Е —Б –і—А—Г–≥–Є–Љ–Є —А–∞—Б—В–µ–љ–Є—П–Љ–Є —Б–Є–љ–Є–є —Б–≤–µ—В —В–∞–Ї–ґ–µ –≤—Л–Ј—Л–≤–∞–ї —В–Њ—А–Љ–Њ–ґ–µ–љ–Є–µ —А–Њ—Б—В–∞ —Б—В–µ–±–ї—П –Є –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤. –Э–∞ —А–Є—Б, 7 –њ–Њ–Ї–∞–Ј–∞–љ—Л —А–∞—Б—В–µ–љ–Є—П –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞, –≤—Л—А–∞—Й–µ–љ–љ—Л–µ –љ–∞ —Б–Є–љ–µ–Љ –Є –љ–∞ –Ї—А–∞—Б–љ–Њ–Љ —Б–≤–µ—В—Г. –†–∞—Б—В–µ–љ–Є—П –љ–∞ –Ї—А–∞—Б–љ–Њ–Љ —Б–≤–µ—В—Г –Є–Љ–µ–ї–Є –њ–Њ—З—В–Є –≤ 3 —А–∞–Ј–∞ –±–Њ–ї—М—И—Г—О –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤ - 79 –і–Љ2 –њ–Њ —Б—А–∞–≤–љ–µ–љ–Є—О —Б –≤–∞—А–Є–∞–љ—В–Њ–Љ –љ–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г (27 –і–Љ2). –С–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј, –љ–∞–±–ї—О–і–∞–µ–Љ—Л–є –Њ–±—Л—З–љ–Њ –љ–∞ —Б–Є–љ–µ–Љ —Б–≤–µ—В—Г, –љ–µ –Љ–Њ–≥ –Ї–Њ–Љ–њ–µ–љ—Б–Є—А–Њ–≤–∞—В—М –і–µ—Д–Є—Ж–Є—В –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ —Г —Н—В–Є—Е —А–∞—Б—В–µ–љ–Є–є, —З—В–Њ –њ—А–Є–≤–Њ–і–Є–ї–Њ –Ї —Б–љ–Є–ґ–µ–љ–Є—О –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є. –£ —А–∞—Б—В–µ–љ–Є–є –њ–Њ–і—Б–Њ–ї–љ–µ—З–љ–Є–Ї–∞ –љ–∞'–Ї—А–∞—Б–љ–Њ–Љ —Б–≤–µ—В—Г –љ–∞–±–ї—О–і–∞–ї—Б—П –Є–Ј–±—Л—В–Њ—З–љ—Л–є «–±–∞–ї–ї–∞—Б—В–љ—Л–є» —А–Њ—Б—В –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В—М–µ–≤ –≤ —Г—Й–µ—А–± —А–Њ—Б—В—Г —А–µ–њ—А–Њ–і—Г–Ї—В–Є–≤–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤.
–Ш—В–∞–Ї, –Ї—А–∞—Б–љ—Л–є —Б–≤–µ—В —Б –Љ–∞–Ї—Б–Є–Љ—Г–Љ–Њ–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П 640-670 –љ–Љ —Б–њ–Њ—Б–Њ–±—Б—В–≤—Г–µ—В –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–Љ—Г —А–Њ—Б—В—Г –ї–Є—Б—В—М–µ–≤ –Є –Њ—Б–µ–≤—Л—Е –Њ—А–≥–∞–љ–Њ–≤. –Э–∞ –Ї—А–∞—Б–љ–Њ–Љ —Б–≤–µ—В—Г –Љ—Л –љ–∞–±–ї—О–і–∞–ї–Є —Б–∞–Љ—Л–є –≤—Л—Б–Њ–Ї–Є–є —А–Њ—Б—В–Њ–≤–Њ–є —Н—Д—Д–µ–Ї—В, —Б—В–Є–Љ—Г–ї–Є—А–Њ–≤–∞–љ–љ—Л–є, –≤–µ—А–Њ—П—В–љ–Њ, –Є —Д–Њ—В–Њ—А–µ—Ж–µ–њ—В–Њ—А–Њ–Љ –Ї—А–∞—Б–љ–Њ–≥–Њ —Б–≤–µ—В–∞ - —Д–Є—В–Њ—Е—А–Њ–Љ–Њ–Љ.
–°–Є–љ–Є–є —Б–≤–µ—В —В–Њ—А–Љ–Њ–Ј–Є—В —А–Њ—Б—В —Б—В–µ–±–ї—П –Є –њ–ї–Њ—Й–∞–і—М –ї–Є—Б—В—М–µ–≤, —З—В–Њ –њ—А–Є–≤–Њ–і–Є—В –Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О —А–∞—Б—В–µ–љ–Є–є —Б –љ–Є–Ј–Ї–Њ–є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М—О. –Т –Ј–µ–ї–µ–љ–Њ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞ (–Љ–∞–Ї—Б–Є–Љ—Г–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П 520-550 –љ–Љ) —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М —В–Њ–љ–Ї–Є–µ –ї–Є—Б—В—М—П —Б –Љ–µ–љ—М—И–Є–Љ —З–Є—Б–ї–Њ–Љ –Ї–ї–µ—В–Њ–Ї –Є —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В–Њ–≤ –Є —Б–∞–Љ—Л–Љ –љ–Є–Ј–Ї–Є–Љ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–Њ–Љ –љ–∞ –µ–і–Є–љ–Є—Ж—Г –њ–ї–Њ—Й–∞–і–Є –ї–Є—Б—В–∞, –љ–Њ —Б–∞–Љ—Л–Љ –≤—Л—Б–Њ–Ї–Є–Љ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–Њ–Љ –≤ —А–∞—Б—З–µ—В–µ –љ–∞ —Е–ї–Њ—А–Њ–њ–ї–∞—Б—В; –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–є –±—Л–ї–∞ –љ–Є–Ј–Ї–Њ–є.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ» –Ї–∞–ґ–і–∞—П –Є–Ј —В—А–µ—Е –Њ—Б–љ–Њ–≤–љ—Л—Е –Њ–±–ї–∞—Б—В–µ–є –§–Р–† (—Б–Є–љ—П—П, –Ј–µ–ї–µ–љ–∞—П –Є –Ї—А–∞—Б–љ–∞—П), –≤–Ј—П—В–∞—П –≤ –Њ—В–і–µ–ї—М–љ–Њ—Б—В–Є, –Љ–∞–ї–Њ –њ—А–Є–≥–Њ–і–љ–∞ –і–ї—П –≤—Л—А–∞—Й–Є–≤–∞–љ–Є—П —А–∞—Б—В–µ–љ–Є–є –Є —В–Њ–ї—М–Ї–Њ –Є–Ј–ї—Г—З–µ–љ–Є–µ, –≤–Ј—П—В–Њ–µ –≤ –Њ–њ—А–µ–і–µ–ї–µ–љ–љ–Њ–Љ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–Є —Н–љ–µ—А–≥–Є–Є –њ–Њ –≤—Б–µ–Љ—Г —Б–њ–µ–Ї—В—А—Г, –Љ–Њ–ґ–µ—В –Њ–±–µ—Б–њ–µ—З–Є—В—М –≤—Л—А–∞—Й–Є–≤–∞–љ–Є–µ –њ–Њ–ї–љ–Њ—Ж–µ–љ–љ—Л—Е —А–∞—Б—В–µ–љ–Є–є. –°–Є–ї—М–љ–Њ–µ –љ–∞—А—Г—И–µ–љ–Є–µ —Н—В–Њ–≥–Њ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є—П, –љ–∞–њ—А–Є–Љ–µ—А, –Ї–Њ–≥–і–∞ —А–∞—Б—В–µ–љ–Є—П –њ–Њ–ї—Г—З–∞—О—В –Љ–∞–Ї—Б–Є–Љ—Г–Љ –Є–Ј–ї—Г—З–µ–љ–Є—П —В–Њ–ї—М–Ї–Њ –≤ —Б–Є–љ–µ–є –Њ–±–ї–∞—Б—В–Є —Б–њ–µ–Ї—В—А–∞, –њ—А–Є–≤–Њ–і–Є—В –Ї —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є—О –љ–Є–Ј–Ї–Њ—А–Њ—Б–ї—Л—Е —А–∞—Б—В–µ–љ–Є–є —Б –≤—Л—Б–Њ–Ї–Є–Љ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–Њ–Љ, –љ–Њ –љ–Є–Ј–Ї–Њ–є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М—О. –°–Є–ї—М–љ—Л–є «–њ–µ—А–µ–Ї–Њ—Б» –њ–Њ —Б–њ–µ–Ї—В—А—Г –≤ –Ї—А–∞—Б–љ—Г—О –Њ–±–ї–∞—Б—В—М —Б–њ–µ–Ї—В—А–∞, –љ–∞–Њ–±–Њ—А–Њ—В, –њ—А–Є–≤–Њ–і–Є—В –Ї –Є–Ј–ї–Є—И–љ–µ–Љ—Г —А–Њ—Б—В—Г –≤–µ–≥–µ—В–∞—В–Є–≤–љ—Л—Е –Њ—А–≥–∞–љ–Њ–≤ –≤ —Г—Й–µ—А–± –≥–µ–љ–µ—А–∞—В–Є–≤–љ—Л–Љ.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л–є —Б–Њ—Б—В–∞–≤ —Б–≤–µ—В–∞, —В–∞–Ї –ґ–µ –Ї–∞–Ї –Є –µ–≥–Њ –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В—М, —П–≤–ї—П–µ—В—Б—П —Б–Є–ї—М–љ—Л–Љ –Љ–Њ—А—Д–Њ–≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–Љ —Д–∞–Ї—В–Њ—А–Њ–Љ, —А–µ–≥—Г–ї–Є—А—Г—О—Й–Є–Љ –Ї–∞–Ї —А–Њ—Б—В–Њ–≤—Л–µ, —В–∞–Ї –Є —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Є–µ —А–µ–∞–Ї—Ж–Є–Є –≤ —Б–Є—Б—В–µ–Љ–µ —Ж–µ–ї–Њ–≥–Њ —А–∞—Б—В–µ–љ–Є—П. –Я—А–Є —Н—В–Њ–Љ –ґ–µ–ї–∞—В–µ–ї—М–љ–Њ –Є–Љ–µ—В—М —Б–ї–µ–і—Г—О—Й–µ–µ —Б–Њ–Њ—В–љ–Њ—И–µ–љ–Є–µ —Н–љ–µ—А–≥–Є–Є –њ–Њ —Б–њ–µ–Ї—В—А—Г –§–Р–† –≤ —А–∞—Б—В–µ–љ–Є–µ–≤–Њ–і—З–µ—Б–Ї–Є—Е –ї–∞–Љ–њ–∞—Е –і–ї—П –Љ–љ–Њ–≥–Є—Е —Б–µ–ї—М—Б–Ї–Њ—Е–Њ–Ј—П–є—Б—В–≤–µ–љ–љ—Л—Е —А–∞—Б—В–µ–љ–Є–є: 25-30%-–≤ —Б–Є–љ–µ–є –Њ–±–ї–∞—Б—В–Є (380-490 –љ–Љ), 20% –≤ –Ј–µ–ї–µ–љ–Њ–є (490-590 –љ–Љ) –Є 50% - –≤ –Ї—А–∞—Б–љ–Њ–є –Њ–±–ї–∞—Б—В–Є (600-700 –љ–Љ).
–Э–Є–ґ–µ –њ—А–Є–≤–Њ–і—П—В—Б—П –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ—Л–µ –њ–Њ–Ї–∞–Ј–∞—В–µ–ї–Є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є —А–∞—Б—В–µ–љ–Є–є, –њ–Њ–ї—Г—З–∞–µ–Љ—Л–µ –≤ —Г—Б–ї–Њ–≤–Є—П—Е —Д–Є—В–Њ—В—А–Њ–љ–∞ –њ—А–Є –Њ–њ—В–Є–Љ–∞–ї—М–љ—Л—Е —Б–≤–µ—В–Њ–≤—Л—Е —А–µ–ґ–Є–Љ–∞—Е (—Б —Б–Њ–±–ї—О–і–µ–љ–Є–µ–Љ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ –і—А—Г–≥–Є—Е —Д–∞–Ї—В–Њ—А–Њ–≤). –Т —Н—В–Є—Е —Г—Б–ї–Њ–≤–Є—П—Е —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –њ–Њ–і–і–µ—А–ґ–Є–≤–∞–ї—Б—П –≤ —В–µ—З–µ–љ–Є–µ –≤—Б–µ–≥–Њ —Б–≤–µ—В–Њ–≤–Њ–≥–Њ –і–љ—П –±–µ–Ј –њ–Њ–ї—Г–і–µ–љ–љ—Л—Е –і–µ–њ—А–µ—Б—Б–Є–є, —Б—Г—В–Њ—З–љ–Њ–µ —Г—Б–≤–Њ–µ–љ–Є–µ –°–Ю2 –±—Л–ї–Њ –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –≤—Л—И–µ, —З–µ–Љ –њ—А–Є —Б–Њ–ї–љ–µ—З–љ–Њ–Љ –Њ—Б–≤–µ—Й–µ–љ–Є–Є, —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є —А–Њ—Б—В –±—Л–ї–Є —Е–Њ—А–Њ—И–Њ —Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞–љ—Л. –Т –Є—В–Њ–≥–µ –њ–Њ–ї—Г—З–µ–љ—Л —Г—А–Њ–ґ–∞–Є —Б 1 –Љ2 –њ–ї–Њ—Й–∞–і–Є –Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ –±–Њ–ї–µ–µ –≤—Л—Б–Њ–Ї–Є–µ, —З–µ–Љ –≤ –њ–Њ–ї–µ, –њ—А–Є —Б–Њ–Ї—А–∞—Й–µ–љ–љ–Њ–Љ –≤–µ–≥–µ—В–∞—Ж–Є–Њ–љ–љ–Њ–Љ –њ–µ—А–Є–Њ–і–µ: –њ—И–µ–љ–Є—Ж–∞ - 2,2- 2,7 –Ї–≥ –Ј–µ—А–љ–∞ –Ј–∞ 75 –і–љ–µ–є (–Њ—В –њ–Њ—Б–µ–≤–∞), –Њ–≥—Г—А—Ж—Л - 24 –Ї–≥ –њ–ї–Њ–і–Њ–≤ –Ј–∞ 76 –і–љ–µ–є (–Њ—В –њ–Њ—Б–µ–≤–∞), —В–Њ–Љ–∞—В—Л - 27 –Ї–≥ –њ–ї–Њ–і–Њ–≤ –Ј–∞ 70 –і–љ–µ–є (–Њ—В –њ–Њ—Б–µ–≤–∞).
–†–µ–Ј—Г–ї—М—В–∞—В—Л —Н—В–Њ–є —А–∞–±–Њ—В—Л –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ —А–∞—Б—В–µ–љ–Є—П –Њ–±–ї–∞–і–∞—О—В –±–Њ–ї—М—И–Є–Љ –≥–µ–љ–µ—В–Є—З–µ—Б–Ї–Є–Љ –њ–Њ—В–µ–љ—Ж–Є–∞–ї–Њ–Љ, –Ї–Њ—В–Њ—А—Л–є –Њ–±—Л—З–љ–Њ –љ–µ —А–µ–∞–ї–Є–Ј—Г–µ—В—Б—П –њ–Њ–ї–љ–Њ—Б—В—М—О –≤ –њ–Њ–ї–µ–≤—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е, –≥–і–µ —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є —А–Њ—Б—В —В—А—Г–і–љ–Њ —Б–±–∞–ї–∞–љ—Б–Є—А–Њ–≤–∞—В—М.
–Ы–Є—В–µ—А–∞—В—Г—А–∞
- –Ъ–ї–µ—И–љ–Є–љ –Р. –§., –Ы–µ–±–µ–і–µ–≤–∞ –Х. –Т., –Я—А–Њ—В–∞—Б–Њ–≤–∞ –Э. –Э. –Є –і—А. –Т—Л—А–∞—Й–Є–≤–∞–љ–Є–µ —А–∞—Б—В–µ–љ–Є–є –њ—А–Є –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–Љ –Њ—Б–≤–µ—Й–µ–љ–Є–Є. –Ь.: –°–µ–ї—М—Е–Њ–Ј–≥–Є–Ј, 1959. 128 —Б.
- –Ь–∞—И–Ї–Њ–≤ –С. –°. –Т—Л—А–∞—Й–Є–≤–∞–љ–Є–µ —А–∞—Б—В–µ–љ–Є–є –њ—А–Є –Є—Б–Ї—Г—Б—Б—В–≤–µ–љ–љ–Њ–Љ –Њ—Б–≤–µ—Й–µ–љ–Є–Є. –Ы. –Ъ–Њ–ї–Њ—Б, 1966.287 —Б.
- –Ы–Њ–Љ–∞–љ –Т. –Ь. –Ъ—Г—А—Б —Б–≤–µ—В–Њ–Ї—Г–ї—М—В—Г—А—Л —А–∞—Б—В–µ–љ–Є–є. –Ш–Ј–і. 2-–µ. –Ь: –Т—Л—Б—И. —И–Ї–Њ–ї–∞, 1976. 272 —Б.
- –Э–Є—И–њ–Њ—А–Њ–≤–Є—З –Р. –Р. –§–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–∞—П –і–µ—П—В–µ–ї—М–љ–Њ—Б—В—М —А–∞—Б—В–µ–љ–Є–є –Є –њ—Г—В–Є –њ–Њ–≤—Л—И–µ–љ–Є—П –Є—Е –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є. –Ґ–µ–Њ—А–µ—В–Є—З–µ—Б–Ї–Є–µ –Њ—Б–љ–Њ–≤—Л —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Њ–є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є. –Ь.: –Э–∞—Г–Ї–∞, 1972. –°. 511.
- –Я—А–Њ—В–∞—Б–Њ–≤–∞ –Э. –Э., –Ъ–µ—Д–µ–ї–Є –Т. –Ш., –Ъ–Њ—Д –≠. –Ь. –Є –і—А. –§–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–∞—П –∞–Ї—В–Є–≤–љ–Њ—Б—В—М, —А–Њ—Б—В –Є —Г—А–Њ–≤–µ–љ—М –њ—А–Є—А–Њ–і–љ—Л—Е —А–µ–≥—Г–ї—П—В–Њ—А–Њ–≤ —Г —А–∞—Б—В–µ–љ–Є–є, –≤—Л—А–∞—Й–µ–љ–љ—Л—Е –љ–∞ —Б–≤–µ—В—Г —А–∞–Ј–ї–Є—З–љ–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–Є–Њ—Б—В–Є. –Ґ–µ–Њ—А–µ—В–Є—З–µ—Б–Ї–Є–µ –Њ—Б–љ–Њ–≤—Л —Д–Њ—В–Њ—Б–Є–љ—В–µ—В–Є—З–µ—Б–Ї–Њ–є –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В–Є. –Ь.: –Э–∞—Г–Ї–∞, 1972. –°. 385.
- –Я—А–Њ—В–∞—Б–Њ–≤–∞ –Э. –Э., –Ъ–µ—Д–µ–ї–Є –Т. –Ш. –§–Њ—В–Њ—Б–Є–љ—В–µ–Ј –Є —А–Њ—Б—В –≤—Л—Б—И–Є—Е —А–∞—Б—В–µ–љ–Є–є, –Є—Е –≤–Ј–∞–Є–Љ–Њ—Б–≤—П–Ј—М –Є –Ї–Њ—А—А–µ–ї—П—Ж–Є–Є. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—П —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞. –Ь.: –Э–∞—Г–Ї–∞, 1982. –°. 251.
- –Ъ—Г–њ–µ—А–Љ–∞–љ –Ш. –Р. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Љ–µ—Е–∞–љ–Є–Ј–Љ—Л –∞–і–∞–њ—В–∞—Ж–Є–Є –Є —Г—Б—В–Њ–є—З–Є–≤–Њ—Б—В–Є —А–∞—Б—В–µ–љ–Є–є. –Э–Њ–≤–Њ—Б–Є–±–Є—А—Б–Ї: –Э–∞—Г–Ї–∞, 1972. –°. 5.
- –¶–µ–ї—М–љ–Є–Ї–µ—А –Ѓ. –Ы. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–µ –Њ—Б–љ–Њ–≤—Л —В–µ–љ–µ–≤—Л–љ–Њ—Б–ї–Є–≤–Њ—Б—В–Є –і—А–µ–≤–µ—Б–љ—Л—Е —А–∞—Б—В–µ–љ–Є–є. –Ь.: –Э–∞—Г–Ї–∞, 1978. 212 —Б.
- –Ґ–Є—Е–Њ–Љ–Є—А–Њ–≤ –Р. –Р., –°–Є–і—М–Ї–Њ –§. –ѓ. –°–Њ—Б—В–Њ—П–љ–Є–µ –њ–Є–≥–Љ–µ–љ—В–љ–Њ–≥–Њ –∞–њ–њ–∞—А–∞—В–∞ –Є —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–µ —Б—В—А—Г–Ї—В—Г—А—Л —Ж–µ–љ–Њ–Ј–Њ–≤ —А–µ–і–Є—Б–∞ –≤ —Б–≤—П–Ј–Є —Б –Є—Е –њ—А–Њ–і—Г–Ї—В–Є–≤–љ–Њ—Б—В—М—О –њ—А–Є —А–∞–Ј–ї–Є—З–љ–Њ–є –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ—Б—В–Є –Є —Б–њ–µ–Ї—В—А–µ –Є–Ј–ї—Г—З–µ–љ–Є—П. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—П —А–∞—Б—В–µ–љ–Є–є. 1982. –Ґ. 29. –Т—Л–њ. 3. –°. 457.
- –Ь–Њ–Ї—А–Њ–љ–Њ—Б–Њ–≤ –Р. –Ґ. –≠–љ–і–Њ–≥–µ–љ–љ–∞—П —А–µ–≥—Г–ї—П—Ж–Є—П —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј–∞ –≤ —Ж–µ–ї–Њ–Љ —А–∞—Б—В–µ–љ–Є–є. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—П —А–∞—Б—В–µ–љ–Є–є. 1978. –Ґ. 25. –Т—Л–њ. 5. –°. 938.
- –Ъ–µ—Д–µ–ї–Є –Т. –Ш. –Я—А–Є—А–Њ–і–љ—Л–µ –Є–љ–≥–Є–±–Є—В–Њ—А—Л —А–Њ—Б—В–∞ –Є —Д–Є—В–Њ–≥–Њ—А–Љ–Њ–љ—Л. –Ь.: –Э–∞—Г–Ї–∞, 1974. 247 —Б.
- –Я—А–Њ—В–∞—Б–Њ–≤–∞ –Э. –Э., –Ы–Њ–ґ–љ–Є–Ї–Њ–≤–∞ –Т, –Э., –Э–Є—З–Є–њ–Њ—А–Њ–≤–Є—З –Р. –Р. –Є –і—А. –†–Њ—Б—В, –∞–Ї—В–Є–≤–љ–Њ—Б—В—М —Д–Є—В–Њ–≥–Њ—А–Љ–Њ–љ–Њ–≤ –Є –Є–љ–≥–Є–±–Є—В–Њ—А–Њ–≤ –Є —Д–Њ—В–Њ—Б–Є–љ—В–µ–Ј —Г –Ї–∞—А–ї–Є–Ї–Њ–≤—Л—Е –Љ—Г—В–∞–љ—В–Њ–≤ –≥–Њ—А–Њ—Е–∞ –≤ —А–∞–Ј–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е —Б–≤–µ—В–Њ–≤–Њ–≥–Њ —А–µ–ґ–Є–Љ–∞. –Ш–Ј–≤. –Р–Э –°–°–°–†. –°–µ—А- –±–Є–Њ–ї., 1980., вДЦ 1. –°. 94.
- –Ь—Б–°–≥–µ–µ –Ъ. J. The action spectrum, absorplance and quantum yield of photosynthesis in crop plants. Agric. Meteoroi. 1972. V. 9. P. 192.
- –Ъ–∞—А–љ–∞—З—Г–Ї –†. –Р., –Я—А–Њ—В–∞—Б–Њ–≤–∞ –Э. –Э„ –Ф–Њ–±—А–Њ–≤–Њ–ї—М—Б–Ї–Є–є –Ь. –Т. –Є –і—А. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–∞—П –∞–і–∞–њ—В–∞—Ж–Є—П –ї–Є—Б—В–∞ –ї–µ–≤–Ј–µ–Є –Ї —Б–њ–µ–Ї—В—А–∞–ї—М–љ–Њ–Љ—Г —Б–Њ—Б—В–∞–≤—Г —Б–≤–µ—В–∞. –§–Є–Ј–Є–Њ–ї–Њ–≥–Є—П —А–∞—Б—В–µ–љ–Є–є. 1987. –Ґ. 34. –Т—Л–њ. 4. –°. 51.
- –Ґ–Є—Е–Њ–Љ–Є—А–Њ–≤ –Р. –Р., –°–Є–і—М–Ї–Њ –§. –ѓ., –Ы–Є—Б–Њ–≤—Б–Ї–Є–є –Ґ. /VI. –Є –і—А. –Я—А–Њ–±–ї–µ–Љ–∞ –Њ–њ—В–Є–Љ–Є–Ј–∞—Ж–Є–Є —Б–њ–µ–Ї—В—А–∞–ї—М–љ—Л—Е –Є —Н–љ–µ—А–≥–µ—В–Є—З–µ—Б–Ї–Є—Е —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї –Є–Ј–ї—Г—З–µ–љ–Є—П —А–∞—Б—В–µ–љ–Є–µ–≤–Њ–і—З–µ—Б–Ї–Є—Е –ї–∞–Љ–њ. –Ъ—А–∞—Б–љ–Њ—П—А—Б–Ї. –†–Њ—В–∞–њ—А–Є–љ—В. 1983. 47 —Б.
- –Т–Њ—Б–Ї—А–µ—Б–µ–љ—Б–Ї–∞—П –Э. –Я. –§–Њ—В–Њ—А–µ–≥—Г–ї—П—В–Њ—А–љ—Л–µ –∞—Б–њ–µ–Ї—В—Л –Љ–µ—В–∞–±–Њ–ї–Є–Ј–Љ–∞ —А–∞—Б—В–µ–љ–Є–є: 38-–µ –Ґ–Є–Љ–Є—А—П–Ј. —З—В. –Ь: –Э–∞—Г–Ї–∞, 1979. 48 —Б.
–Я—А–Њ—Б–Љ–Њ—В—А–Њ–≤: 28991
–Ф–∞—В–∞: –Я–Њ–љ–µ–і–µ–ї—М–љ–Є–Ї, 26 –°–µ–љ—В—П–±—А—П 2011
–Ю–±—А–∞—В–Є—В–µ –≤–љ–Є–Љ–∞–љ–Є–µ –љ–∞ —Б–ї–µ–і—Г—О—Й–Є–µ —В–Њ–≤–∞—А—Л:
-


-
–Ф–Э–Р–Ґ 600 Green Power - –Т—Л–±–Њ—А –њ—А–Њ—Д–µ—Б—Б–Є–Њ–љ–∞–ї—М–љ—Л—Е —В–µ–њ–ї–Є—З–љ–Є–Ї–Њ–≤
- 5.900 —А—Г–±.
-

-
–Ф–Э–Р–Ґ 400 Green Power -
–°–њ–µ—Ж–Є–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ–љ–∞—П –ї–∞–Љ–њ–∞ –і–ї—П —А–∞—Б—В–µ–љ–Є–є –Ф–Э–Р–Ґ Green Power –Њ—В Philips
- 4.500 —А—Г–±.
-

-
–Ф–Э–∞–Ґ Sylvania GroLux 600 W - –¶–µ–љ–∞-–Ъ–∞—З–µ—Б—В–≤–Њ
- 4.200 —А—Г–±.
-

-
–Ф–Э–∞–Ґ Sylvania GroLux 400 W - –°–њ–µ—Ж–Є–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ–љ–∞—П –ї–∞–Љ–њ–∞ –і–ї—П —А–∞—Б—В–µ–љ–Є–є –Ф–Э–Р–Ґ GroLux –Њ—В Sylvania
- 3.700 —А—Г–±.
-

-
–Ф–Э–∞–Ґ Osram PLANTASTAR 600 W - –°–њ–µ—Ж–Є–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ–љ–∞—П –ї–∞–Љ–њ–∞ –і–ї—П —А–∞—Б—В–µ–љ–Є–є –Ф–Э–Р–Ґ PLANTASTAR –Њ—В Osram
- 4.500 —А—Г–±.
-

-
–Ф–Э–∞–Ґ Osram PLANTASTAR 400 W - –°–њ–µ—Ж–Є–∞–ї–Є–Ј–Є—А–Њ–≤–∞–љ–љ–∞—П –ї–∞–Љ–њ–∞ –і–ї—П —А–∞—Б—В–µ–љ–Є–є –Ф–Э–Р–Ґ PLANTASTAR –Њ—В Osram
- 4.100 —А—Г–±.
 8-920-0234-105
8-920-0234-105
 GrowHobby.ru
GrowHobby.ru
 –Т—Е–Њ–і
–Т—Е–Њ–і